

Endoplasmic reticulum stress in the intestinal epithelium initiates purine metabolite synthesis and promotes Th17 cell differentiation in the gut
- PMID: 36917985
- PMCID: PMC10175221
- DOI: 10.1016/j.immuni.2023.02.018
Endoplasmic reticulum stress in the intestinal epithelium initiates purine metabolite synthesis and promotes Th17 cell differentiation in the gut
Abstract
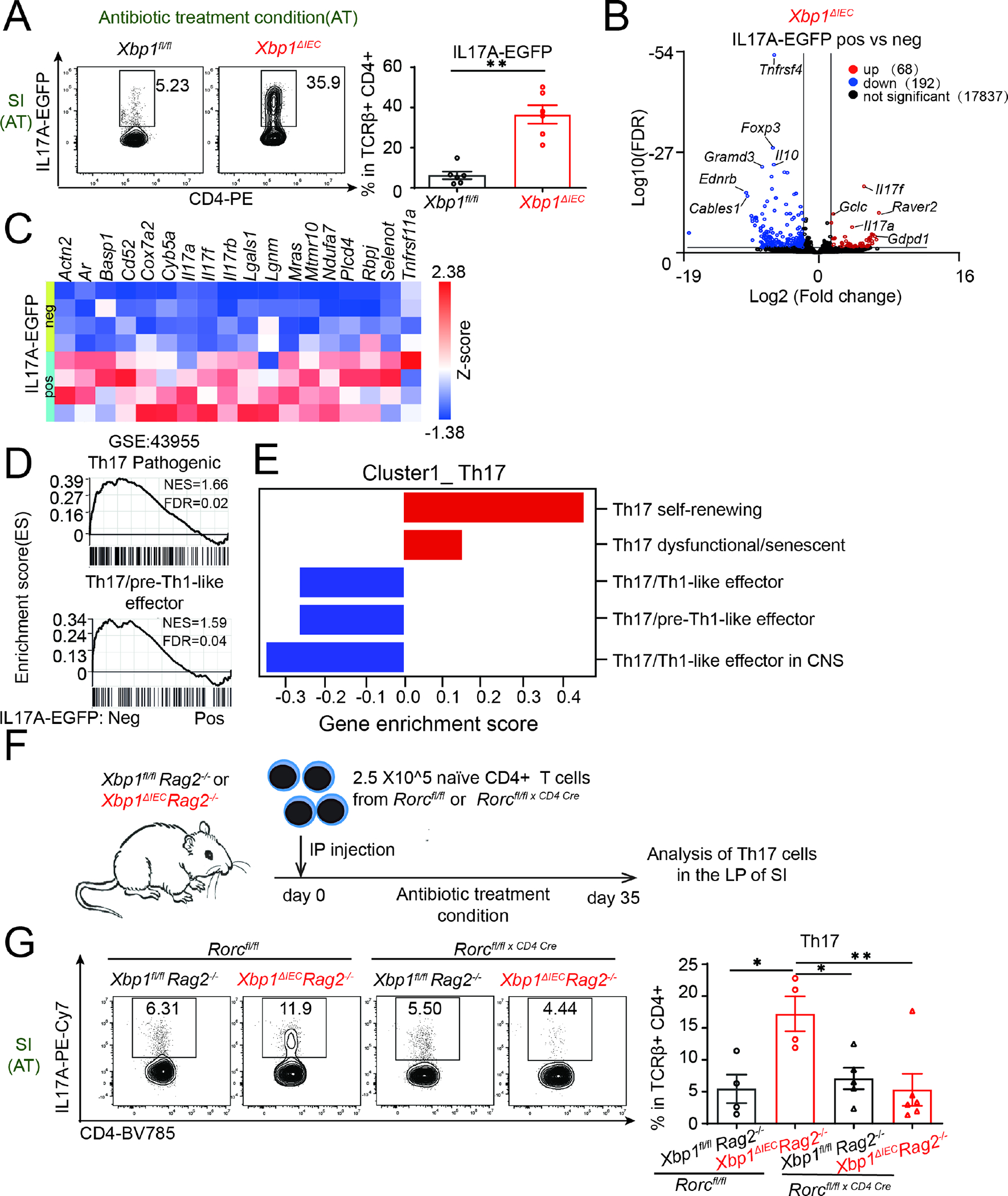
Intestinal IL-17-producing T helper (Th17) cells are dependent on adherent microbes in the gut for their development. However, how microbial adherence to intestinal epithelial cells (IECs) promotes Th17 cell differentiation remains enigmatic. Here, we found that Th17 cell-inducing gut bacteria generated an unfolded protein response (UPR) in IECs. Furthermore, subtilase cytotoxin expression or genetic removal of X-box binding protein 1 (Xbp1) in IECs caused a UPR and increased Th17 cells, even in antibiotic-treated or germ-free conditions. Mechanistically, UPR activation in IECs enhanced their production of both reactive oxygen species (ROS) and purine metabolites. Treating mice with N-acetyl-cysteine or allopurinol to reduce ROS production and xanthine, respectively, decreased Th17 cells that were associated with an elevated UPR. Th17-related genes also correlated with ER stress and the UPR in humans with inflammatory bowel disease. Overall, we identify a mechanism of intestinal Th17 cell differentiation that emerges from an IEC-associated UPR.
Keywords: Citrobacter rodentium; ROS signals; TH17 cells; commensal bacterial; epithelial endoplasmic reticulum stress; inflammatory bowel disease; purine metabolism.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures






Comment in
-
Cellular stress and TH17 cell induction in the gut.Nat Rev Gastroenterol Hepatol. 2023 Jun;20(6):344. doi: 10.1038/s41575-023-00781-4. Nat Rev Gastroenterol Hepatol. 2023. PMID: 37020084 No abstract available.
References
-
- Kaser A, Lee AH, Franke A, Glickman JN, Zeissig S, Tilg H, Nieuwenhuis EE, Higgins DE, Schreiber S, Glimcher LH, and Blumberg RS (2008). XBP1 links ER stress to intestinal inflammation and confers genetic risk for human inflammatory bowel disease. Cell 134, 743–756. 10.1016/j.cell.2008.07.021. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R37 DK044319/DK/NIDDK NIH HHS/United States
- R01 DK088199/DK/NIDDK NIH HHS/United States
- R01 DK051362/DK/NIDDK NIH HHS/United States
- R01 DK053056/DK/NIDDK NIH HHS/United States
- R01 AI169075/AI/NIAID NIH HHS/United States
- 106260/Z/14/Z/WT_/Wellcome Trust/United Kingdom
- R01 DK117565/DK/NIDDK NIH HHS/United States
- WT_/Wellcome Trust/United Kingdom
- 222497/Z/21/Z/WT_/Wellcome Trust/United Kingdom
- R01 DK044319/DK/NIDDK NIH HHS/United States
- R01 DK015070/DK/NIDDK NIH HHS/United States
- R01 AI042347/AI/NIAID NIH HHS/United States
- R37 DK015070/DK/NIDDK NIH HHS/United States
- R01 DK110559/DK/NIDDK NIH HHS/United States
- K12 HD000850/HD/NICHD NIH HHS/United States
- P30 DK034854/DK/NIDDK NIH HHS/United States
- R56 DK053056/DK/NIDDK NIH HHS/United States
- P30 DK020595/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
