

An integrated single cell and spatial transcriptomic map of human white adipose tissue
- PMID: 36922516
- PMCID: PMC10017705
- DOI: 10.1038/s41467-023-36983-2
An integrated single cell and spatial transcriptomic map of human white adipose tissue
Abstract
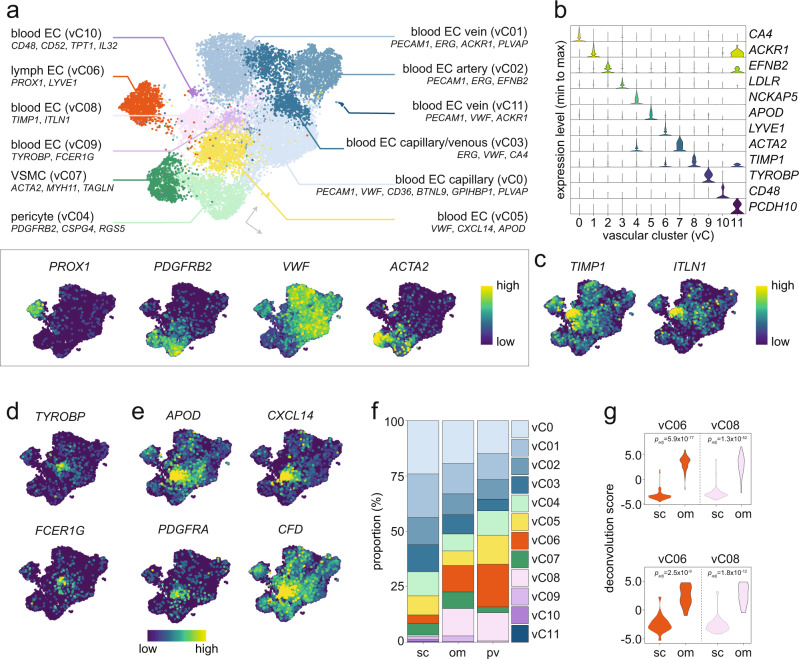
To date, single-cell studies of human white adipose tissue (WAT) have been based on small cohort sizes and no cellular consensus nomenclature exists. Herein, we performed a comprehensive meta-analysis of publicly available and newly generated single-cell, single-nucleus, and spatial transcriptomic results from human subcutaneous, omental, and perivascular WAT. Our high-resolution map is built on data from ten studies and allowed us to robustly identify >60 subpopulations of adipocytes, fibroblast and adipogenic progenitors, vascular, and immune cells. Using these results, we deconvolved spatial and bulk transcriptomic data from nine additional cohorts to provide spatial and clinical dimensions to the map. This identified cell-cell interactions as well as relationships between specific cell subtypes and insulin resistance, dyslipidemia, adipocyte volume, and lipolysis upon long-term weight changes. Altogether, our meta-map provides a rich resource defining the cellular and microarchitectural landscape of human WAT and describes the associations between specific cell types and metabolic states.
© 2023. The Author(s).
Conflict of interest statement
P.L.S. and N.B. are scientific consultants to 10x Genomics, which holds IP rights to the spatial transcriptomics technology. None of the other authors have any conflict of interest to report.
Figures







References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
