

Structural basis of odorant recognition by a human odorant receptor
- PMID: 36922591
- PMCID: PMC10580732
- DOI: 10.1038/s41586-023-05798-y
Structural basis of odorant recognition by a human odorant receptor
Abstract
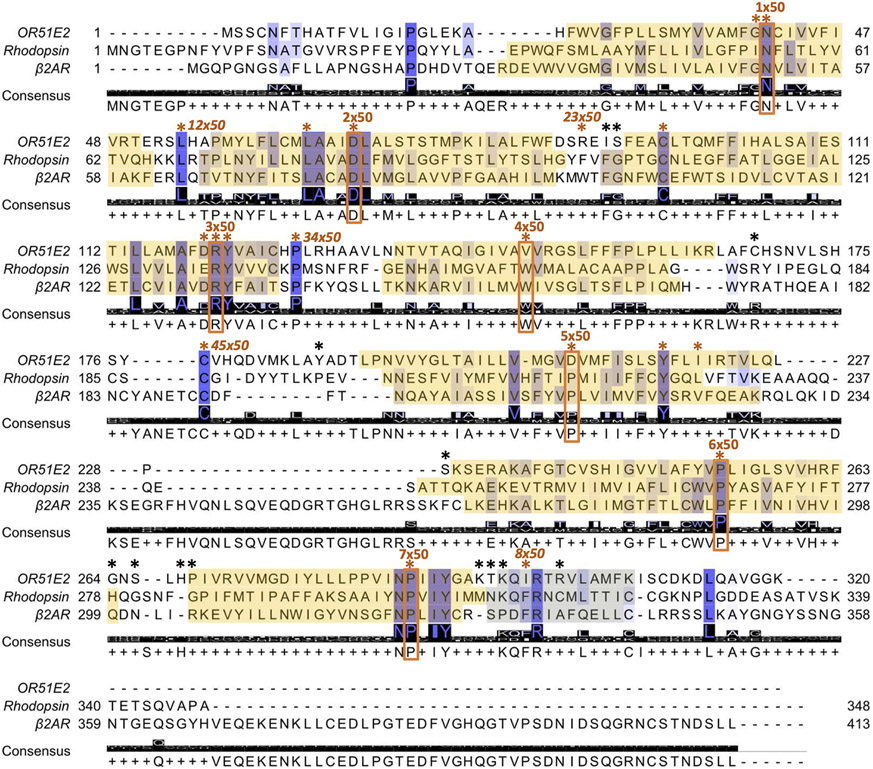
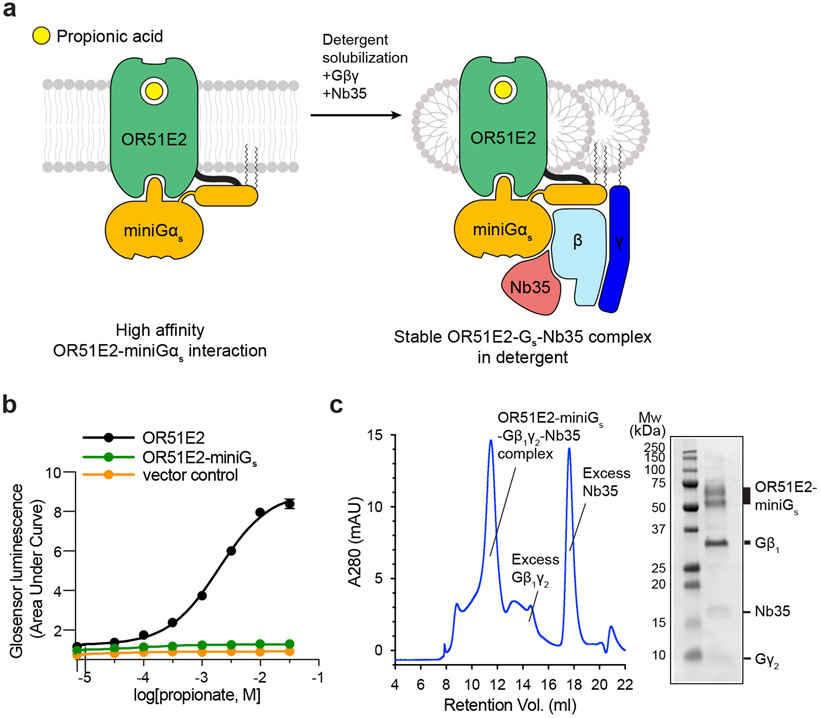
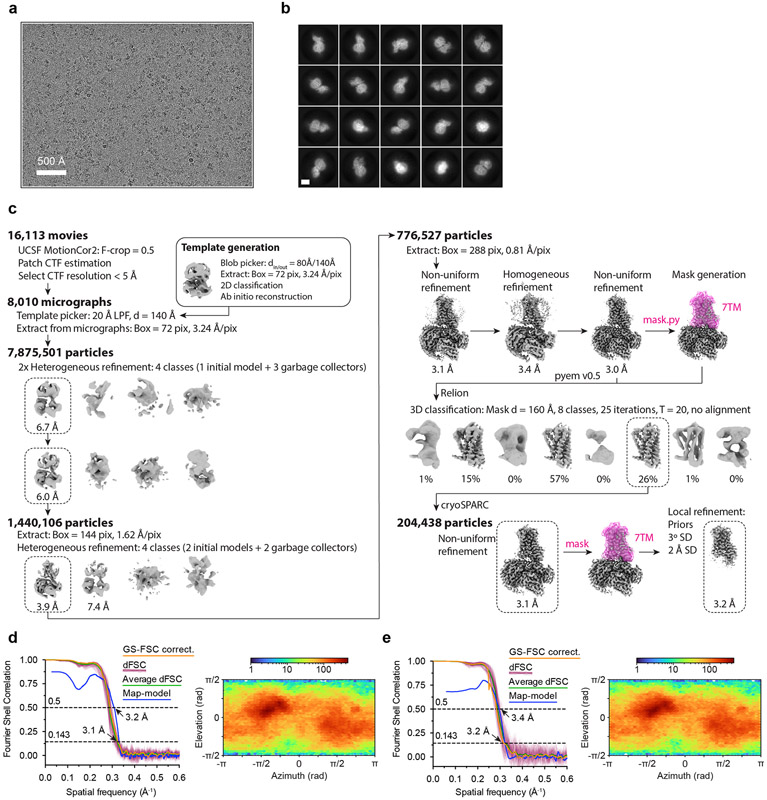
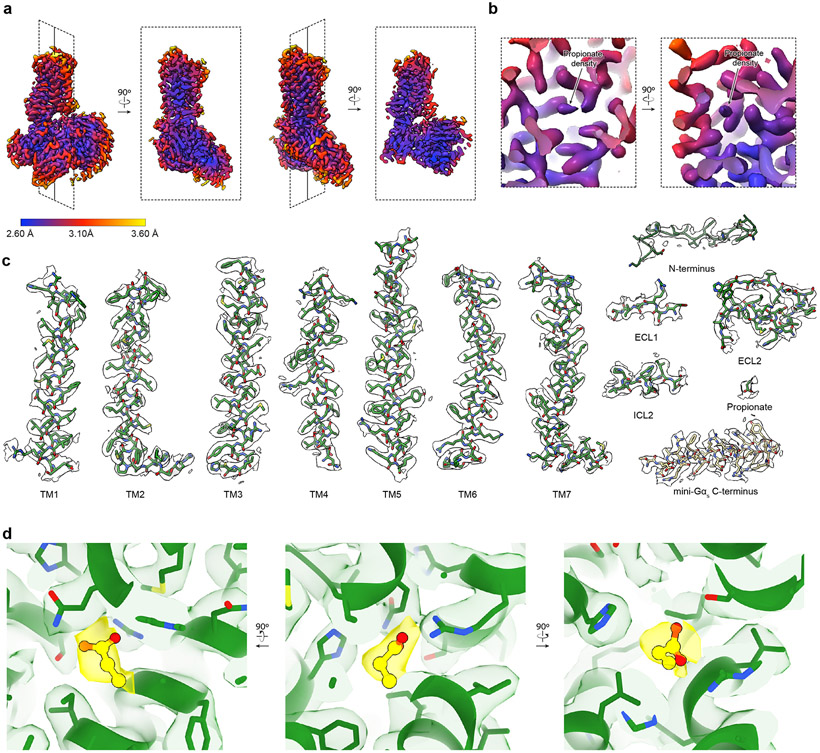
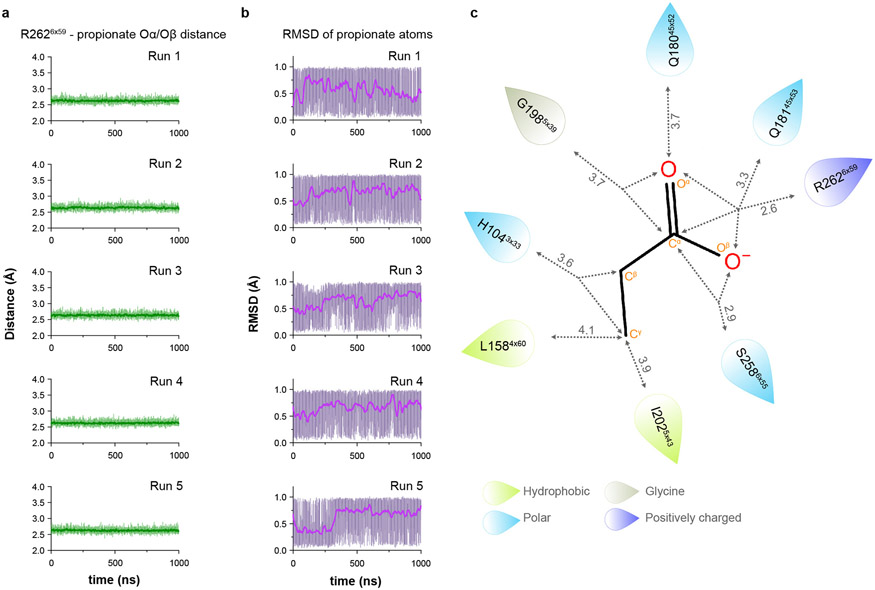
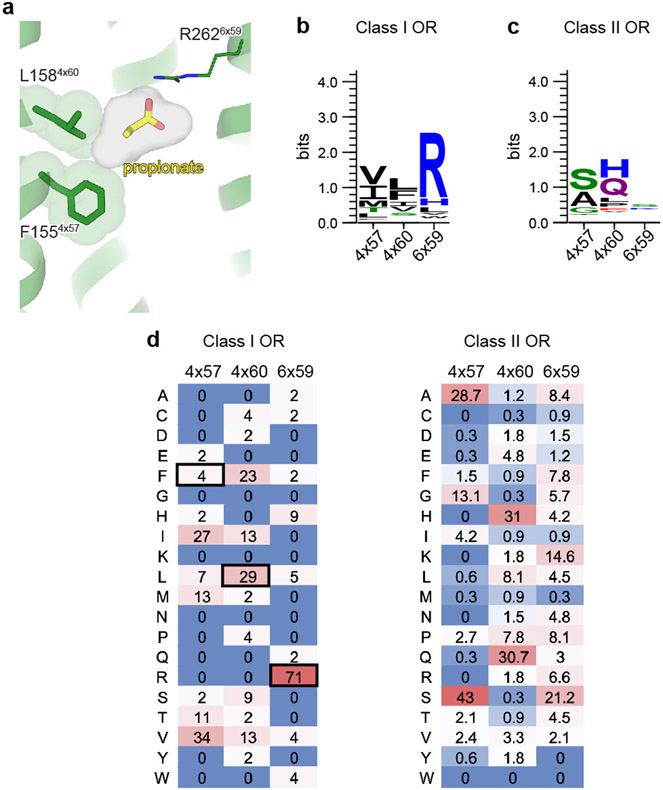
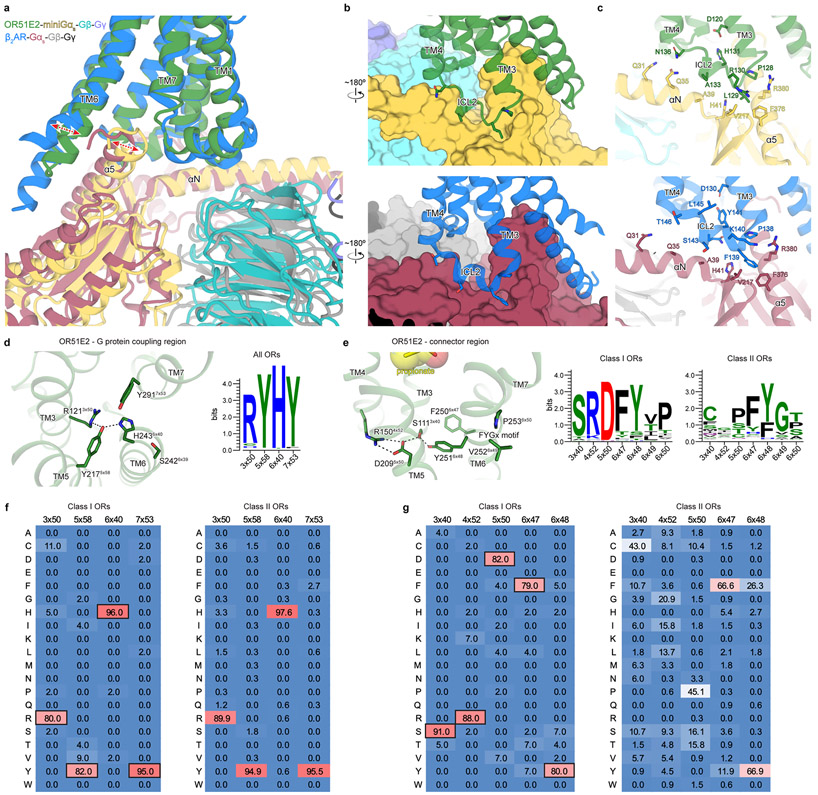
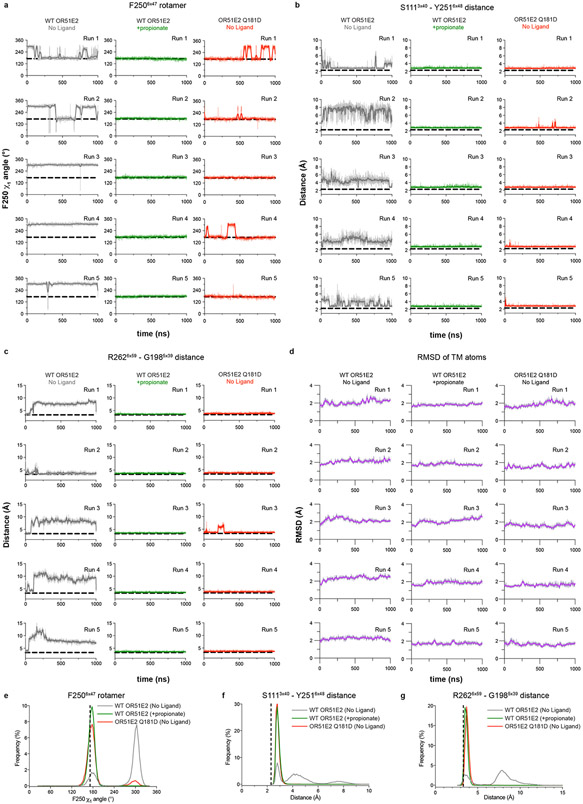
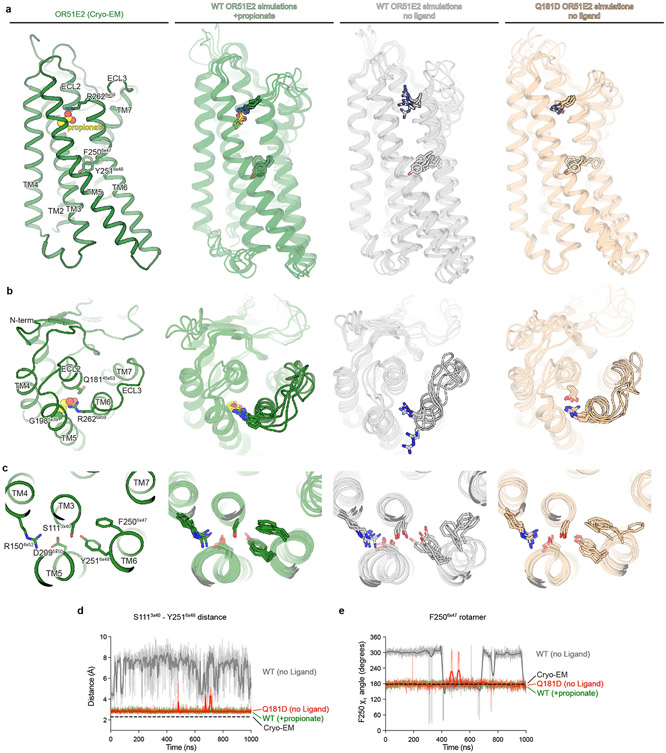
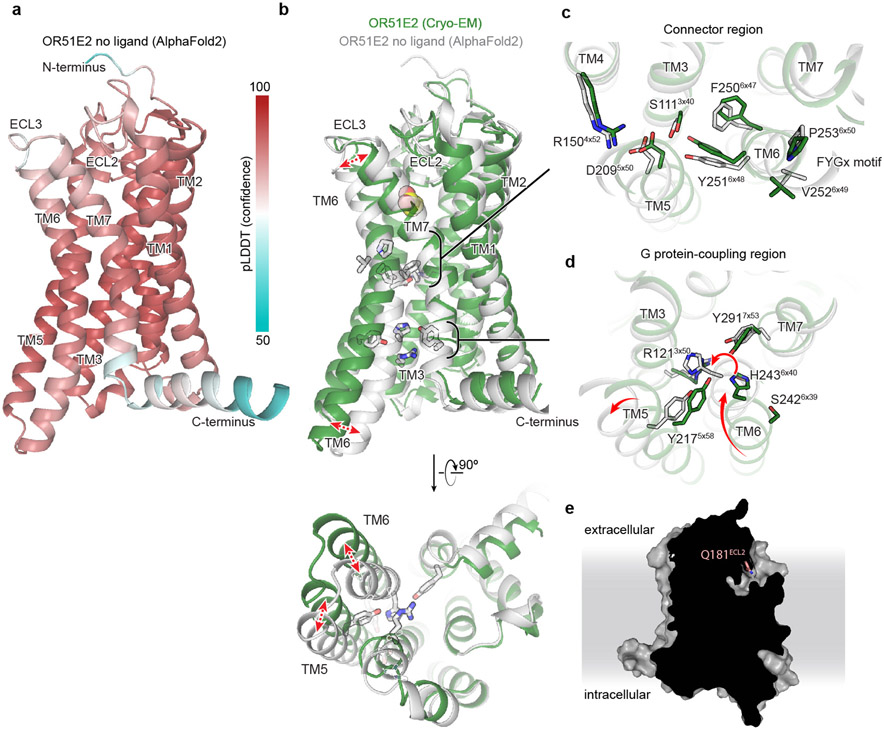
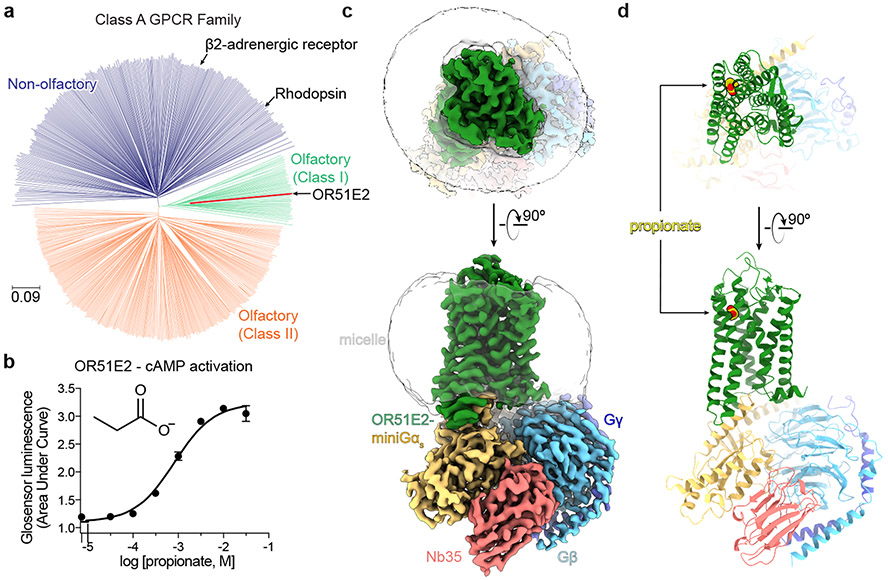
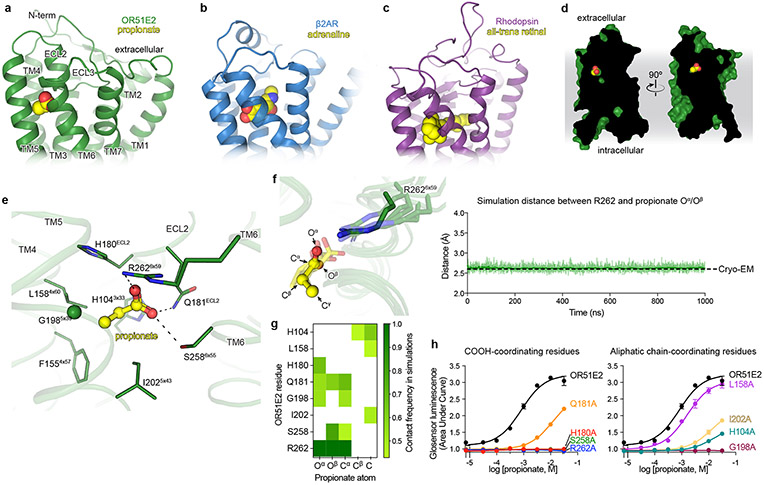
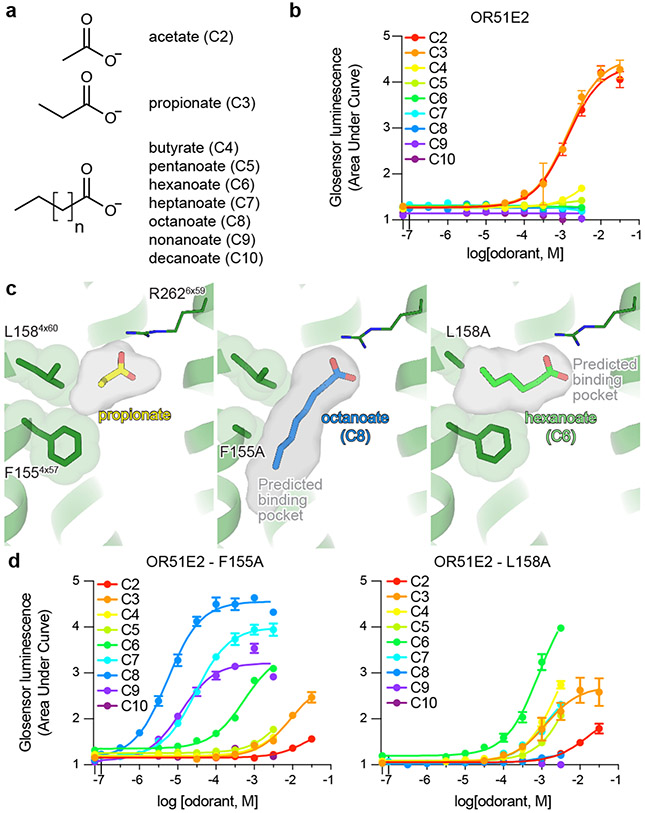
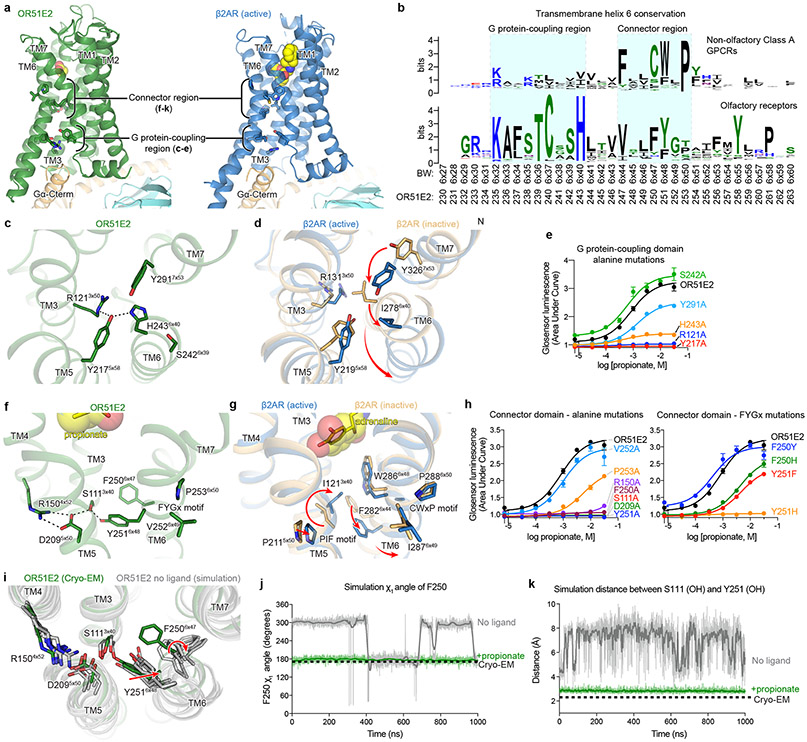
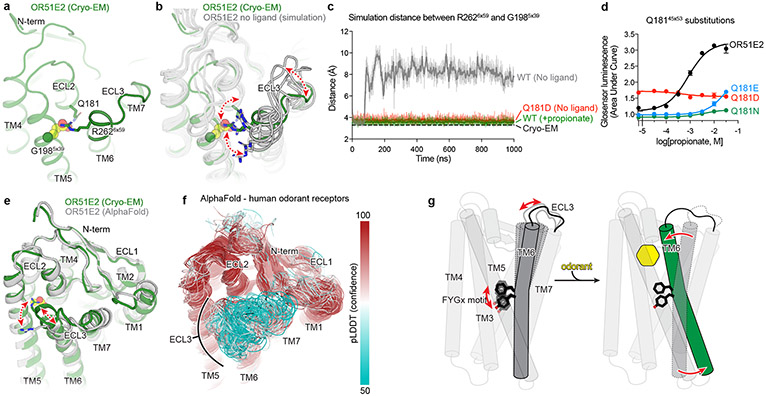
Our sense of smell enables us to navigate a vast space of chemically diverse odour molecules. This task is accomplished by the combinatorial activation of approximately 400 odorant G protein-coupled receptors encoded in the human genome1-3. How odorants are recognized by odorant receptors remains unclear. Here we provide mechanistic insight into how an odorant binds to a human odorant receptor. Using cryo-electron microscopy, we determined the structure of the active human odorant receptor OR51E2 bound to the fatty acid propionate. Propionate is bound within an occluded pocket in OR51E2 and makes specific contacts critical to receptor activation. Mutation of the odorant-binding pocket in OR51E2 alters the recognition spectrum for fatty acids of varying chain length, suggesting that odorant selectivity is controlled by tight packing interactions between an odorant and an odorant receptor. Molecular dynamics simulations demonstrate that propionate-induced conformational changes in extracellular loop 3 activate OR51E2. Together, our studies provide a high-resolution view of chemical recognition of an odorant by a vertebrate odorant receptor, providing insight into how this large family of G protein-coupled receptors enables our olfactory sense.
© 2023. The Author(s), under exclusive licence to Springer Nature Limited.
Figures
References
-
- Buck L, Axel R. A novel multigene family may encode odorant receptors: a molecular basis for odor recognition. Cell. 1991;65(1):175–187. - PubMed
-
- Malnic B, Hirono J, Sato T, Buck LB. Combinatorial receptor codes for odors. Cell. 1999;96(5):713–723. - PubMed
-
- Zhao H, Ivic L, Otaki JM, Hashimoto M, Mikoshiba K, Firestein S. Functional expression of a mammalian odorant receptor. Science. 1998;279(5348):237–242. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
