

Subcellular mRNA localization and local translation of Arhgap11a in radial glial progenitors regulates cortical development
- PMID: 36924763
- PMCID: PMC10132781
- DOI: 10.1016/j.neuron.2023.02.023
Subcellular mRNA localization and local translation of Arhgap11a in radial glial progenitors regulates cortical development
Abstract
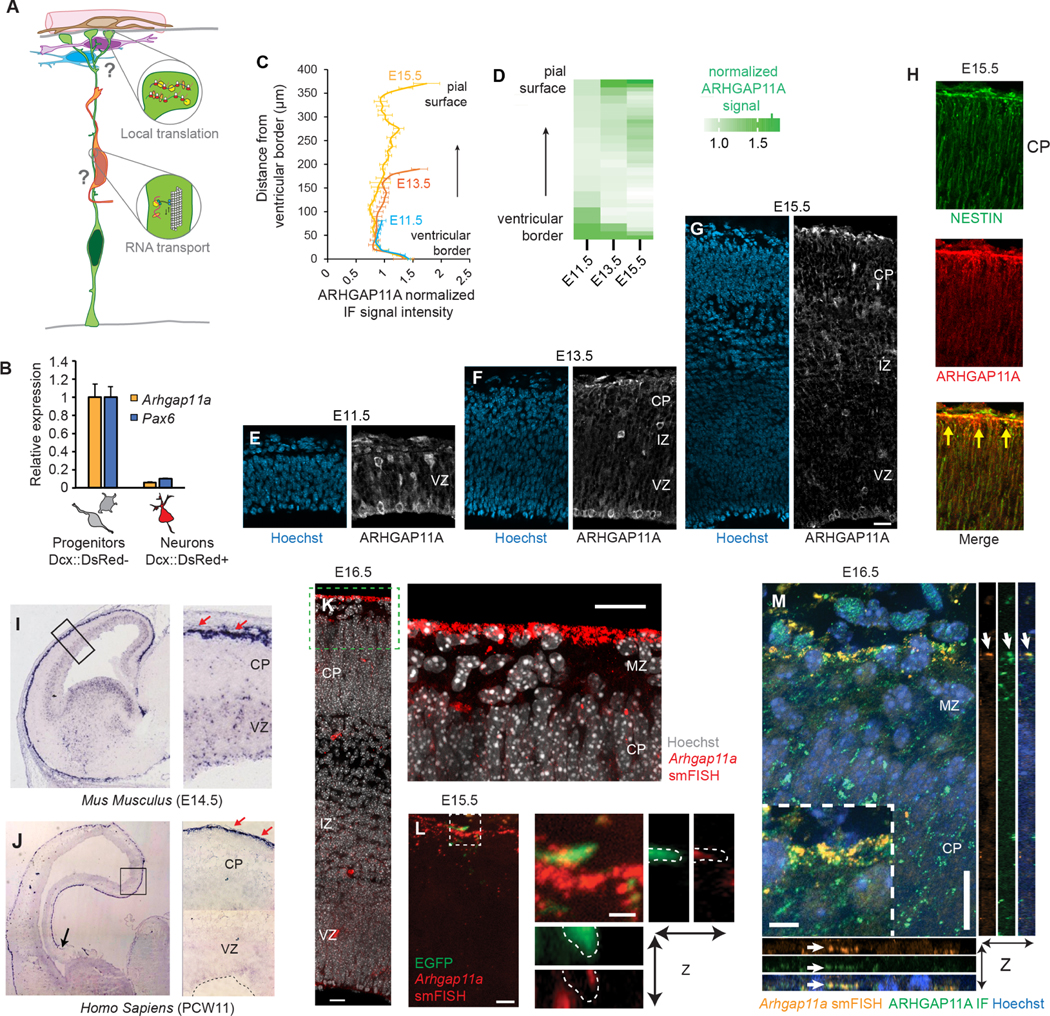
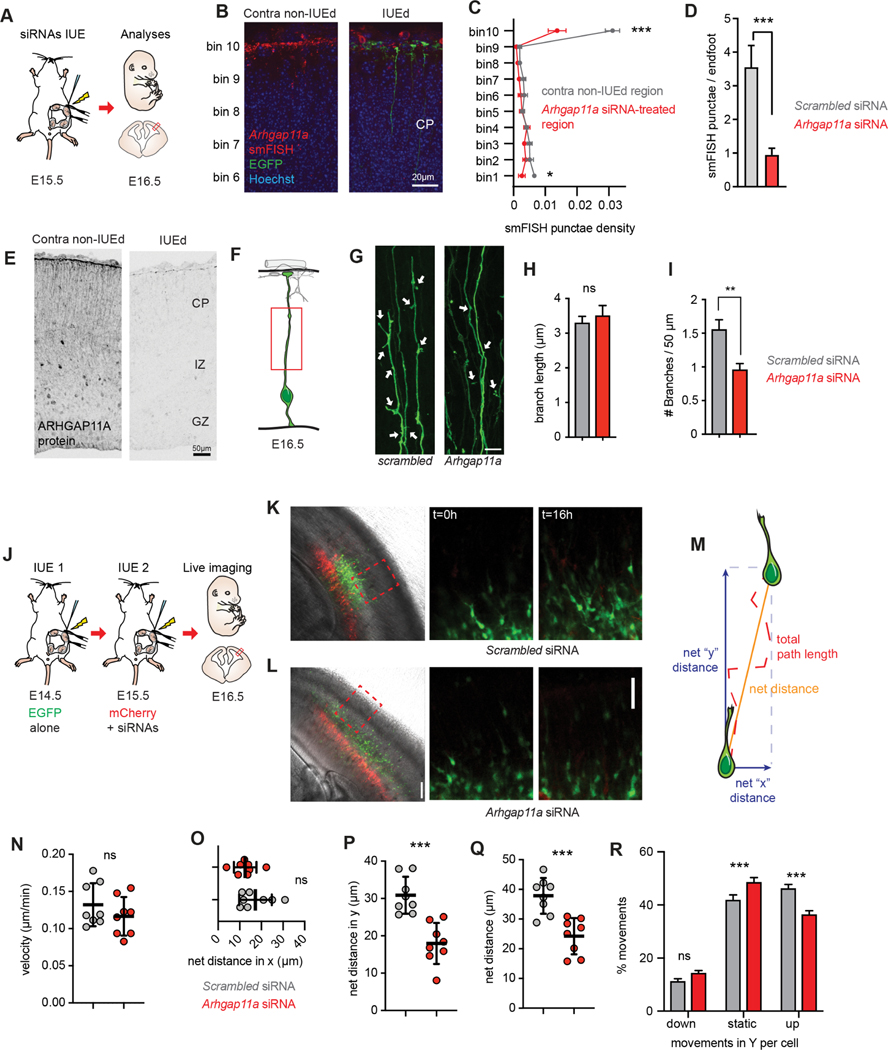
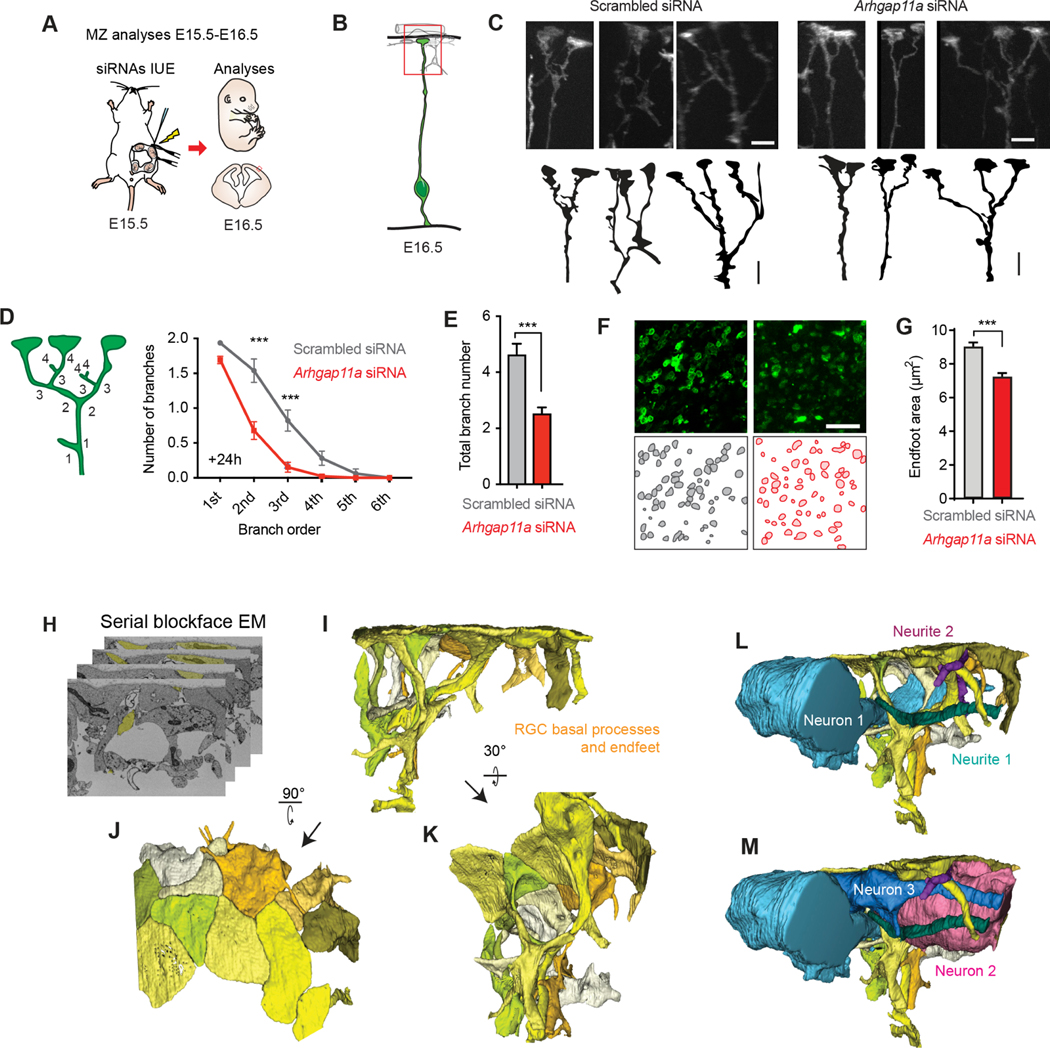
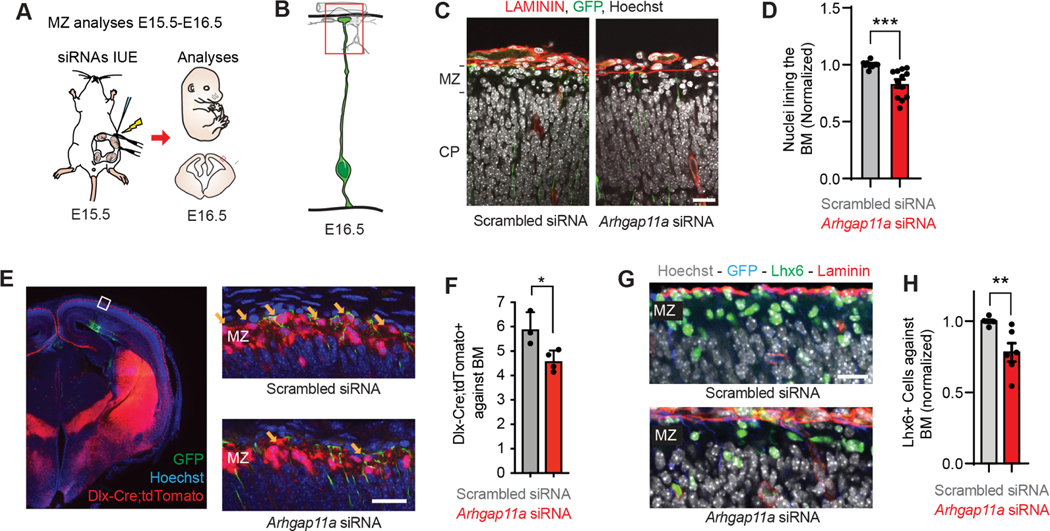
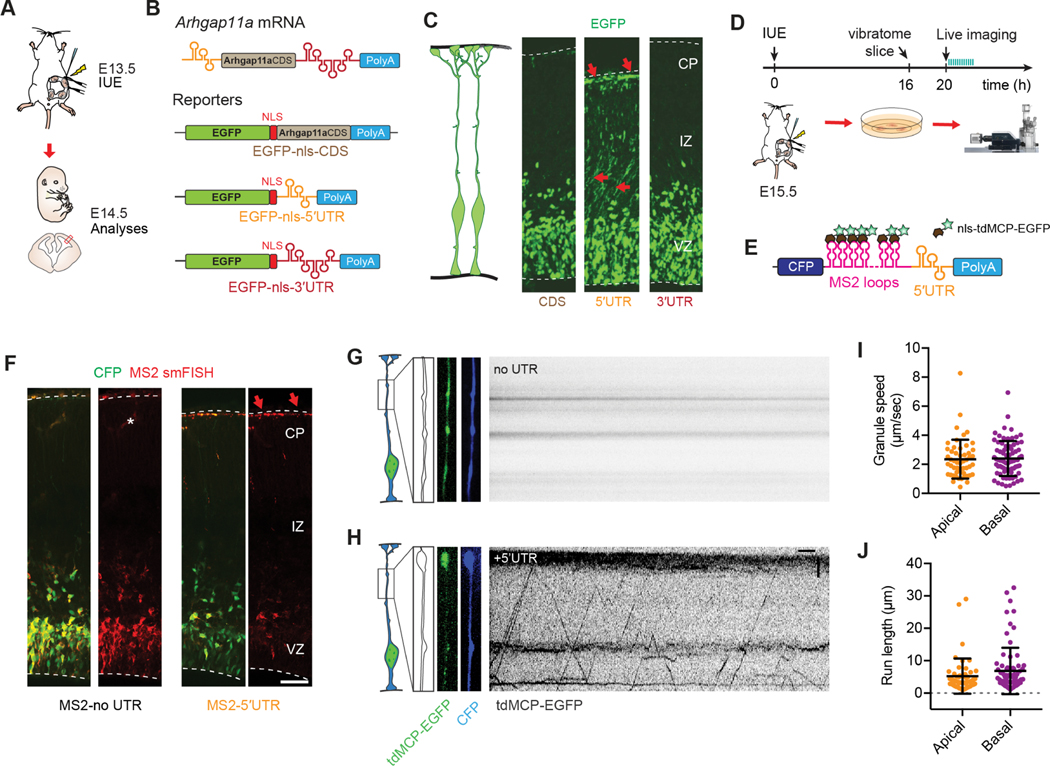
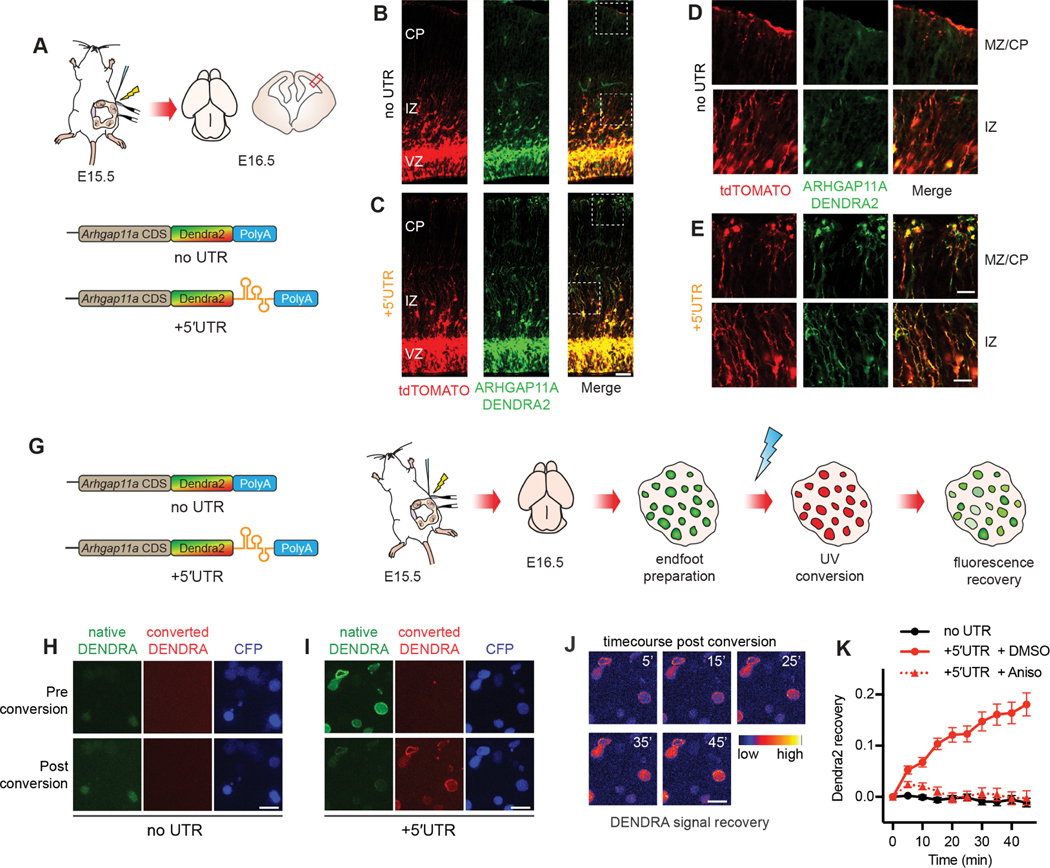
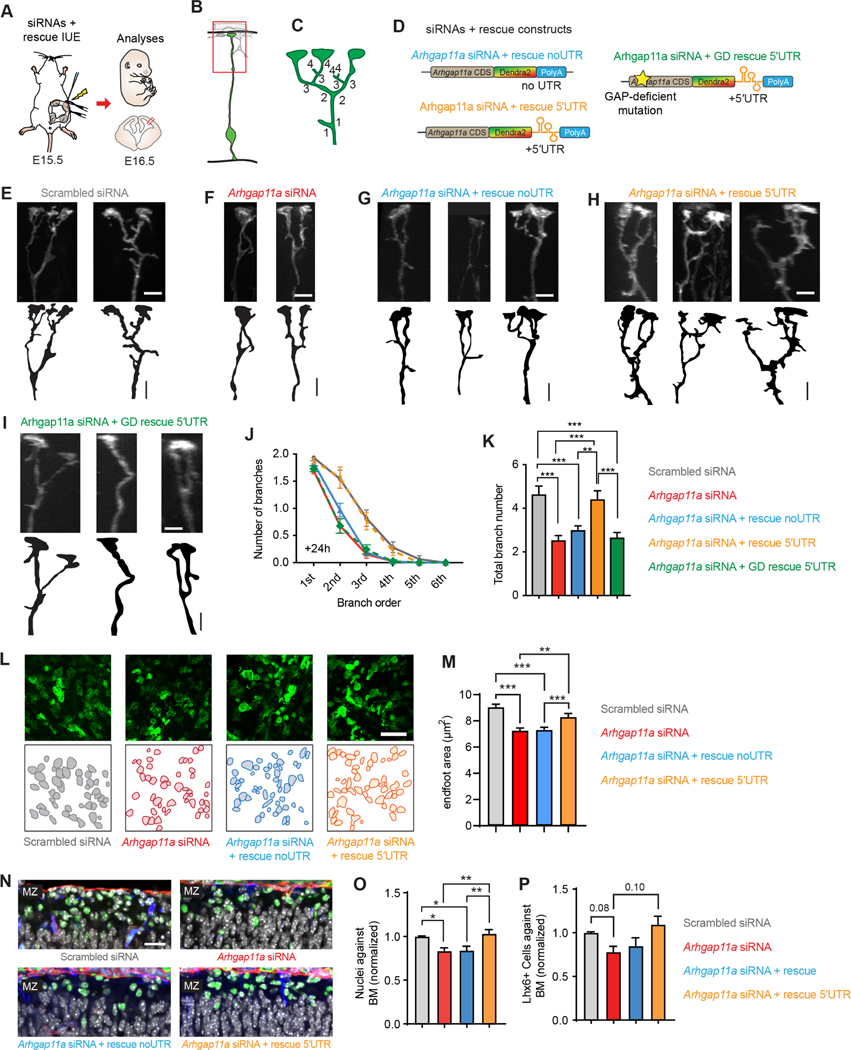
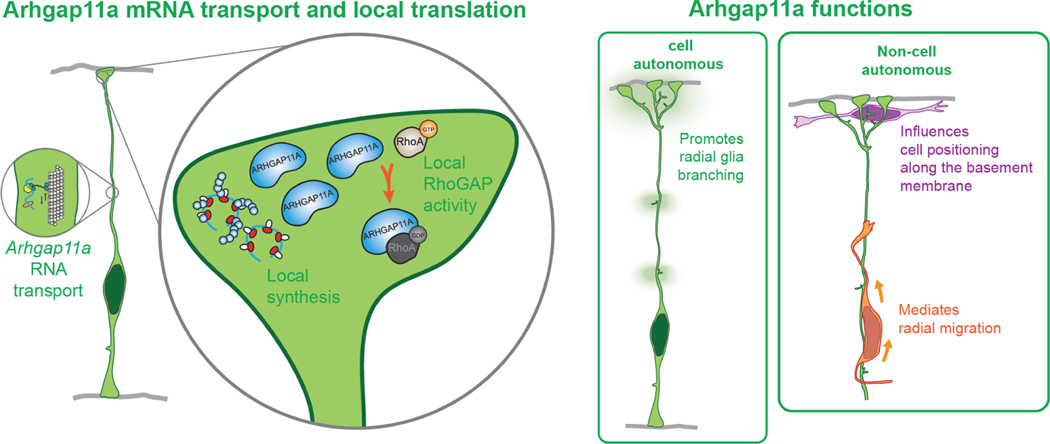
mRNA localization and local translation enable exquisite spatial and temporal control of gene expression, particularly in polarized, elongated cells. These features are especially prominent in radial glial cells (RGCs), which are neural and glial precursors of the developing cerebral cortex and scaffolds for migrating neurons. Yet the mechanisms by which subcellular RGC compartments accomplish their diverse functions are poorly understood. Here, we demonstrate that mRNA localization and local translation of the RhoGAP ARHGAP11A in the basal endfeet of RGCs control their morphology and mediate neuronal positioning. Arhgap11a transcript and protein exhibit conserved localization to RGC basal structures in mice and humans, conferred by the 5' UTR. Proper RGC morphology relies upon active Arhgap11a mRNA transport and localization to the basal endfeet, where ARHGAP11A is locally synthesized. This translation is essential for positioning interneurons at the basement membrane. Thus, local translation spatially and acutely activates Rho signaling in RGCs to compartmentalize neural progenitor functions.
Keywords: Arhgap11a; RhoGAP; cortical development; interneuron positioning neuronal migration; local translation; mRNA localization; mRNA transport; mouse; radial glial endfeet.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Malatesta P, Hartfuss E, and Gotz M. (2000). Isolation of radial glial cells by fluorescent-activated cell sorting reveals a neuronal lineage. Development 127, 5253–5263. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
