

Mechanisms and treatment strategies of demyelinating and dysmyelinating Charcot-Marie-Tooth disease
- PMID: 36926710
- PMCID: PMC10233759
- DOI: 10.4103/1673-5374.367834
Mechanisms and treatment strategies of demyelinating and dysmyelinating Charcot-Marie-Tooth disease
Abstract
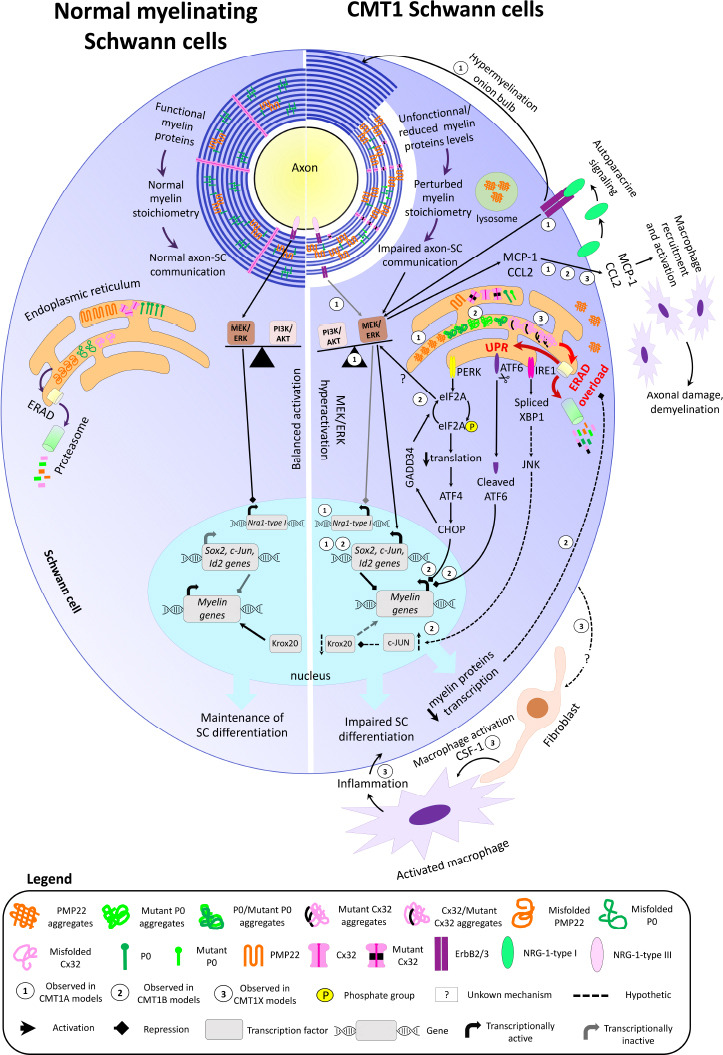
Schwann cells, the myelinating glia of the peripheral nervous system, wrap axons multiple times to build their myelin sheath. Myelin is of paramount importance for axonal integrity and fast axon potential propagation. However, myelin is lacking or dysfunctional in several neuropathies including demyelinating and dysmyelinating Charcot-Marie-Tooth disease. Charcot-Marie-Tooth disease represents the most prevalent inherited neuropathy in humans and is classified either as axonal, demyelinating or dysmyelinating, or as intermediate. The demyelinating or dysmyelinating forms of Charcot-Marie-Tooth disease constitute the majority of the disease cases and are most frequently due to mutations in the three following myelin genes: peripheral myelin protein 22, myelin protein zero and gap junction beta 1 (coding for Connexin 32) causing Charcot-Marie-Tooth disease type 1A, Charcot-Marie-Tooth disease type 1B, and X-linked Charcot-Marie-Tooth disease type 1, respectively. The resulting perturbation of myelin structure and function leads to axonal demyelination or dysmyelination and causes severe disabilities in affected patients. No treatment to cure or slow down the disease progression is currently available on the market, however, scientific discoveries led to a better understanding of the pathomechanisms of the disease and to potential treatment strategies. In this review, we describe the features and molecular mechanisms of the three main demyelinating or dysmyelinating forms of Charcot-Marie-Tooth disease, the rodent models used in research, and the emerging therapeutic approaches to cure or counteract the progression of the disease.
Keywords: Charcot-Marie-Tooth disease; Schwann cells; demyelination and dysmyelination; emerging treatments; endoplasmic reticulum stress; gene therapy; myelin; repair; rodent models; unfolded protein response.
Conflict of interest statement
None
Figures
References
-
- Arthur-Farraj PJ, Latouche M, Wilton DK, Quintes S, Chabrol E, Banerjee A, Woodhoo A, Jenkins B, Rahman M, Turmaine M, Wicher GK, Mitter R, Greensmith L, Behrens A, Raivich G, Mirsky R, Jessen KR. c-Jun reprograms Schwann cells of injured nerves to generate a repair cell essential for regeneration. Neuron. 2012;75:633–647. - PMC - PubMed
-
- Attarian S, Vallat JM, Magy L, Funalot B, Gonnaud PM, Lacour A, Péréon Y, Dubourg O, Pouget J, Micallef J, Franques J, Lefebvre MN, Ghorab K, Al-Moussawi M, Tiffreau V, Preudhomme M, Magot A, Leclair-Visonneau L, Stojkovic T, Bossi L, et al. An exploratory randomised double-blind and placebo-controlled phase 2 study of a combination of baclofen, naltrexone and sorbitol (PXT3003) in patients with Charcot-Marie-Tooth disease type 1A. Orphanet J Rare Dis. 2014;9:199. - PMC - PubMed
-
- Bai Y, Ianokova E, Pu Q, Ghandour K, Levinson R, Martin JJ, Ceuterick-de Groote C, Mazanec R, Seeman P, Shy ME, Li J. Effect of an R69C mutation in the myelin protein zero gene on myelination and ion channel subtypes. Arch Neurol. 2006;63:1787–1794. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous
