

Mechanism of STMN2 cryptic splice-polyadenylation and its correction for TDP-43 proteinopathies
- PMID: 36927019
- PMCID: PMC10148063
- DOI: 10.1126/science.abq5622
Mechanism of STMN2 cryptic splice-polyadenylation and its correction for TDP-43 proteinopathies
Abstract
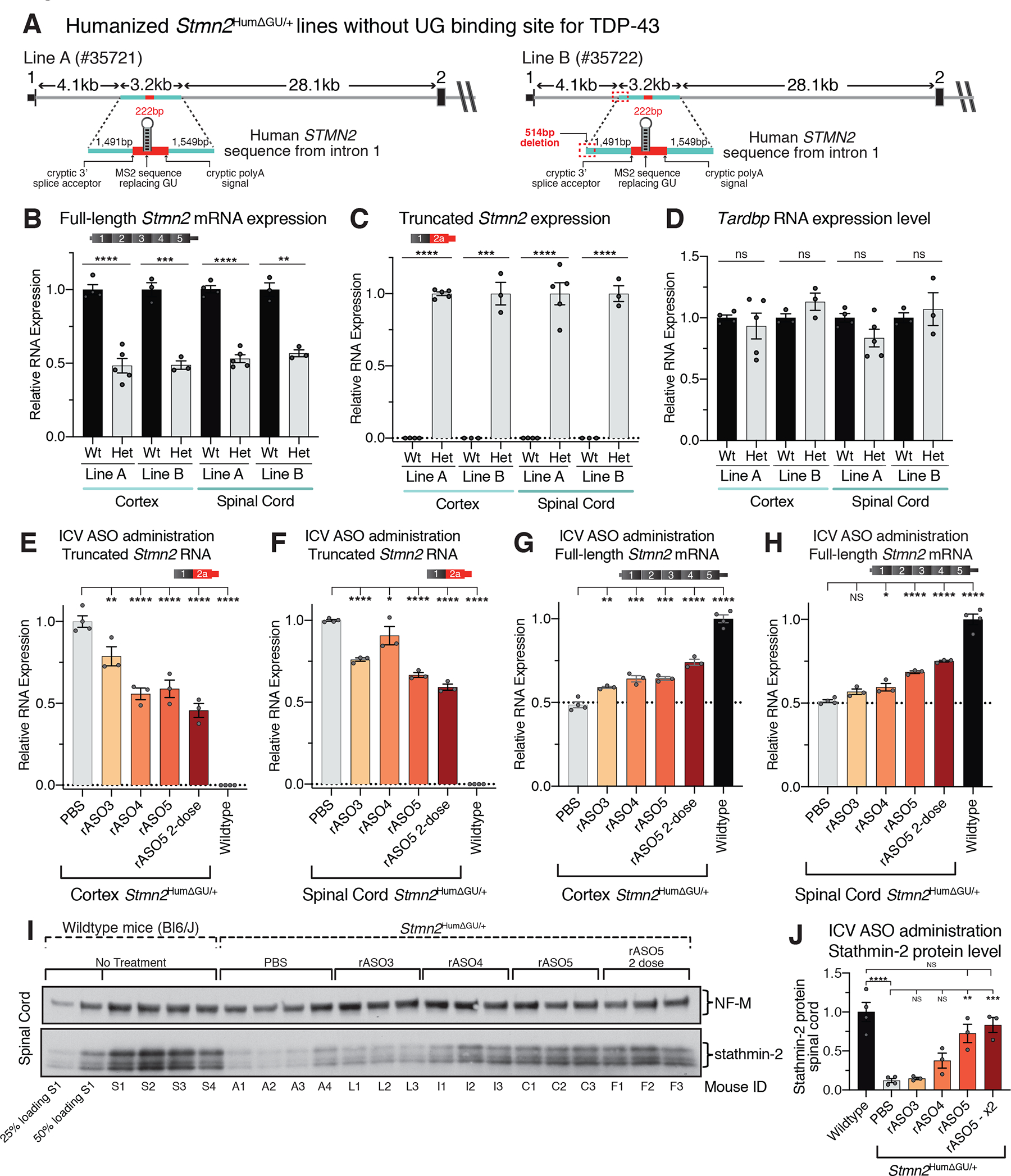
Loss of nuclear TDP-43 is a hallmark of neurodegeneration in TDP-43 proteinopathies, including amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD). TDP-43 mislocalization results in cryptic splicing and polyadenylation of pre-messenger RNAs (pre-mRNAs) encoding stathmin-2 (also known as SCG10), a protein that is required for axonal regeneration. We found that TDP-43 binding to a GU-rich region sterically blocked recognition of the cryptic 3' splice site in STMN2 pre-mRNA. Targeting dCasRx or antisense oligonucleotides (ASOs) suppressed cryptic splicing, which restored axonal regeneration and stathmin-2-dependent lysosome trafficking in TDP-43-deficient human motor neurons. In mice that were gene-edited to contain human STMN2 cryptic splice-polyadenylation sequences, ASO injection into cerebral spinal fluid successfully corrected Stmn2 pre-mRNA misprocessing and restored stathmin-2 expression levels independently of TDP-43 binding.
Conflict of interest statement
Competing interests:
A patent application has been filed describing composition and methods for restoring stathmin-2 expression; publication number US 2021-0252039. D.W.C. and Z.M. are inventors on the patent.
CFB, KL, PJN, FR, are employees of Ionis Pharmaceuticals.
DWC is a consultant for Ionis Pharmaceuticals.
Figures
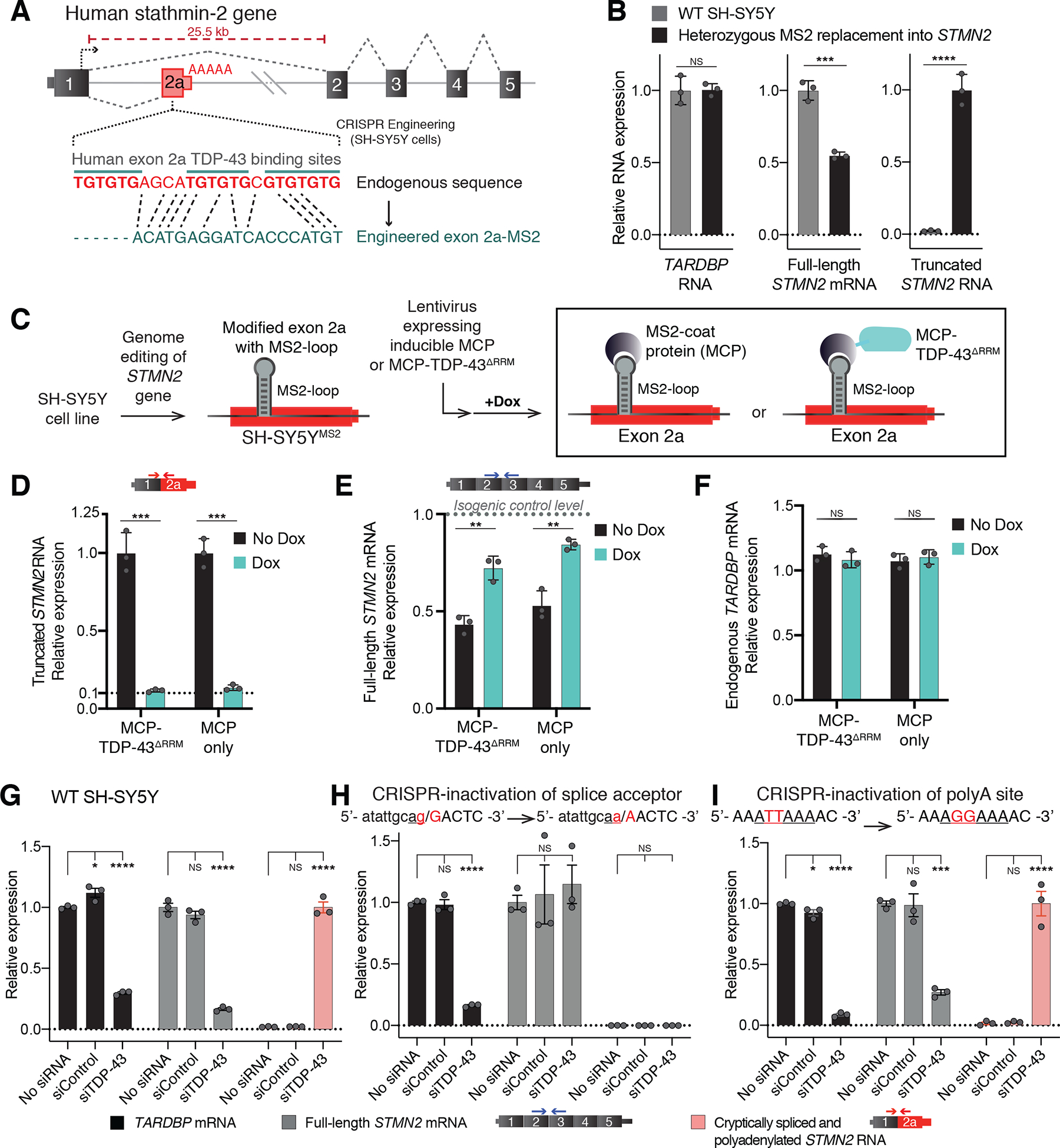
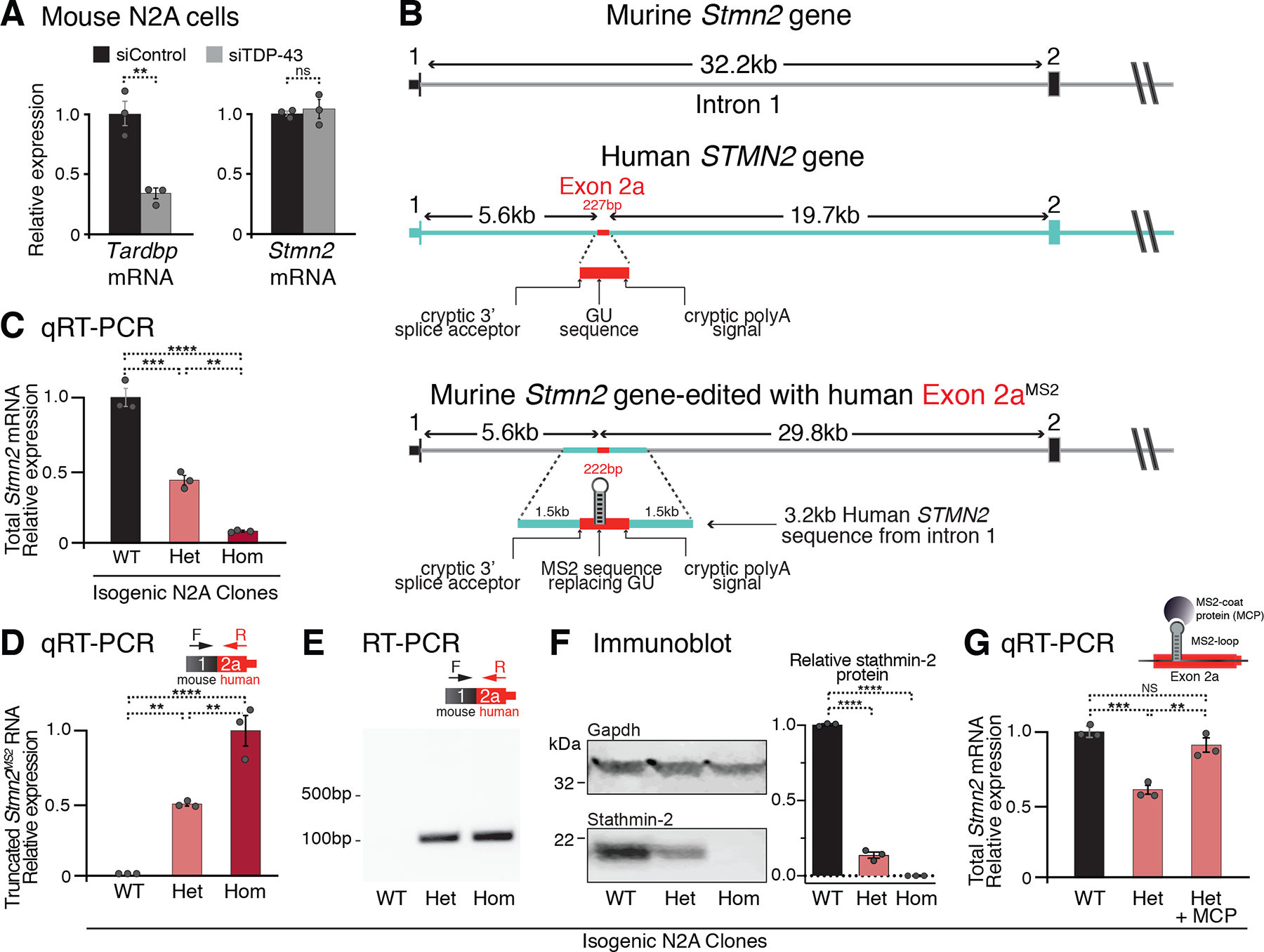
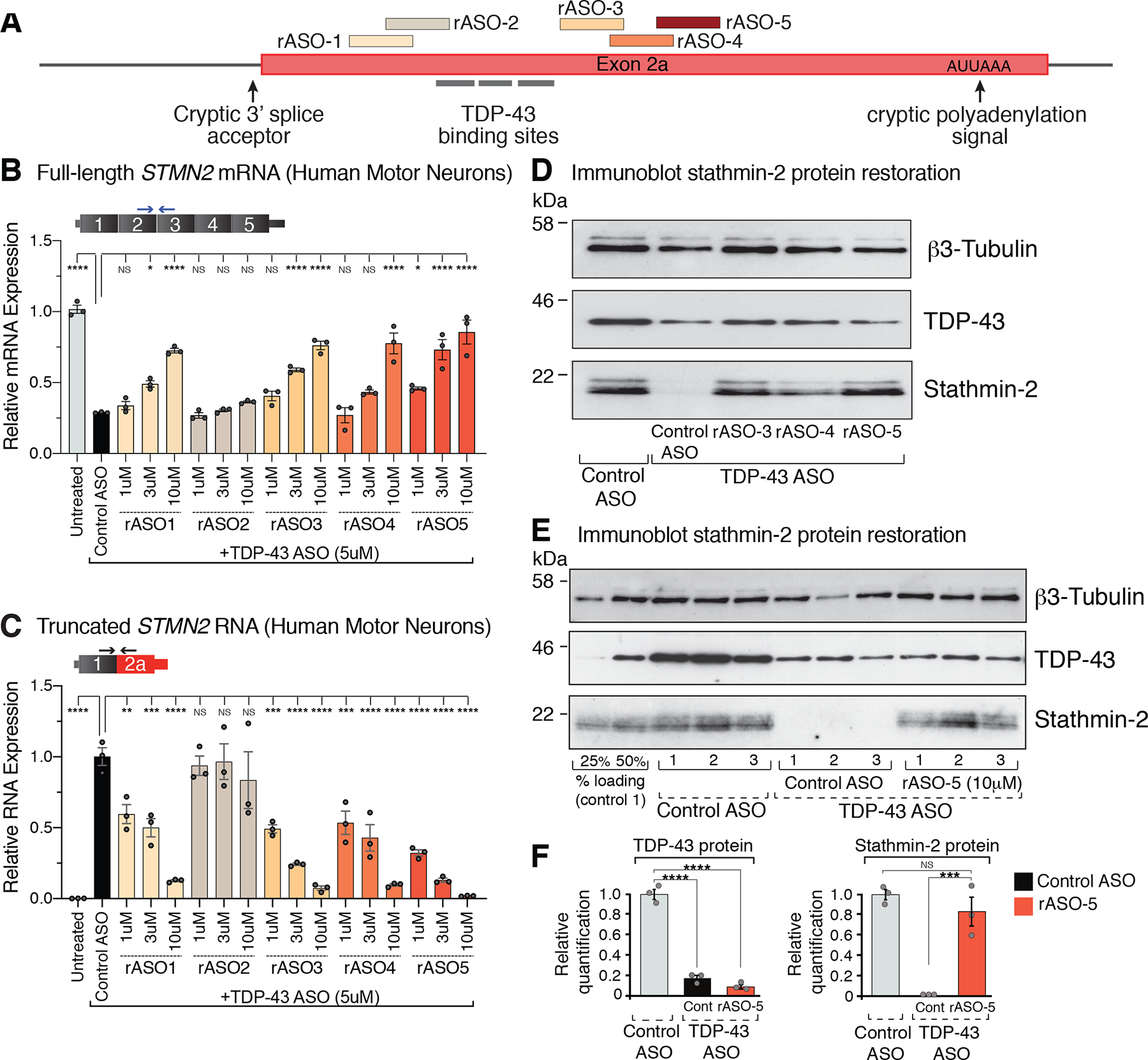
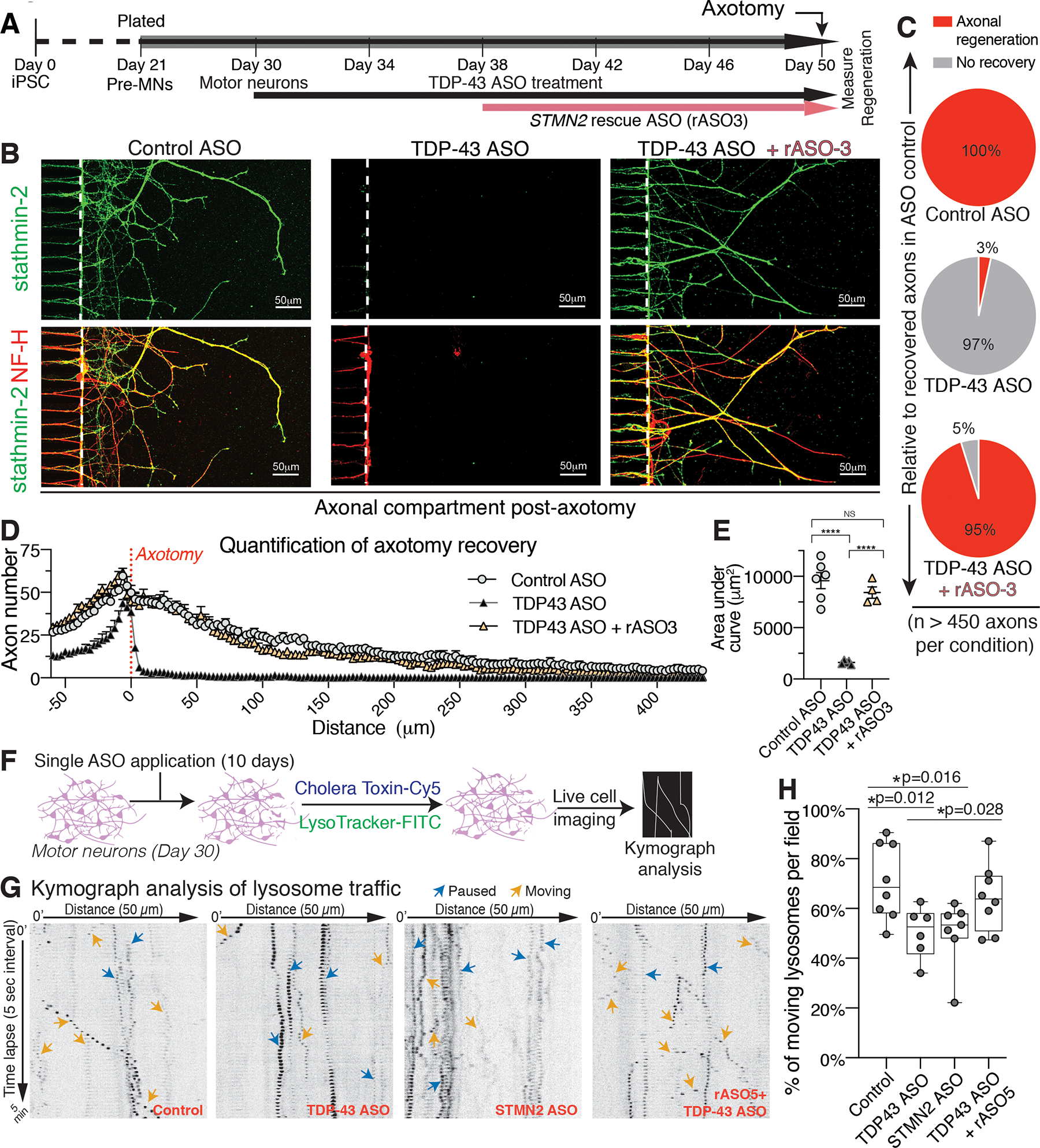
Comment in
-
TDP43 blocks misprocessing of STMN2 RNA.Nat Rev Neurol. 2023 Jun;19(6):326. doi: 10.1038/s41582-023-00806-1. Nat Rev Neurol. 2023. PMID: 37012364 No abstract available.
-
Tackling TDP43 proteinopathies.Nat Rev Drug Discov. 2023 May;22(5):354. doi: 10.1038/d41573-023-00056-2. Nat Rev Drug Discov. 2023. PMID: 37041235 No abstract available.
-
To the rescue by blocking cryptic splicing.Nat Rev Genet. 2023 Jun;24(6):346. doi: 10.1038/s41576-023-00603-y. Nat Rev Genet. 2023. PMID: 37069253 No abstract available.
-
Restoring expression of Stathmin-2: a novel strategy to treat TDP-43 proteinopathies.Signal Transduct Target Ther. 2023 Jul 12;8(1):266. doi: 10.1038/s41392-023-01533-1. Signal Transduct Target Ther. 2023. PMID: 37433765 Free PMC article. No abstract available.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
