

Dps-dependent in vivo mutation enhances long-term host adaptation in Vibrio cholerae
- PMID: 36928244
- PMCID: PMC10104298
- DOI: 10.1371/journal.ppat.1011250
Dps-dependent in vivo mutation enhances long-term host adaptation in Vibrio cholerae
Abstract
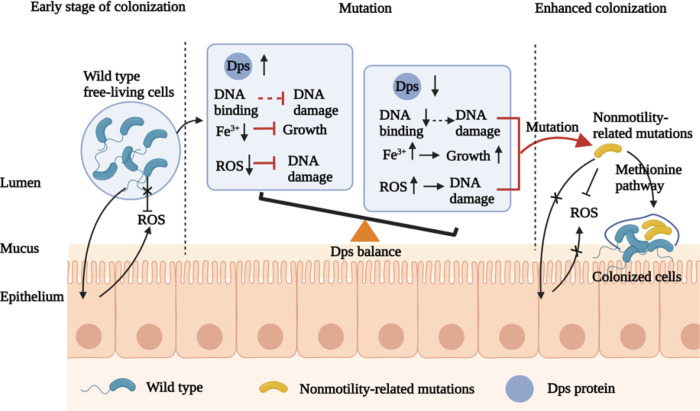
As one of the most successful pathogenic organisms, Vibrio cholerae (V. cholerae) has evolved sophisticated regulatory mechanisms to overcome host stress. During long-term colonization by V. cholerae in adult mice, many spontaneous nonmotile mutants (approximately 10% at the fifth day post-infection) were identified. These mutations occurred primarily in conserved regions of the flagellar regulator genes flrA, flrC, and rpoN, as shown by Sanger and next-generation sequencing, and significantly increased fitness during colonization in adult mice. Intriguingly, instead of key genes in DNA repair systems (mutS, nfo, xthA, uvrA) or ROS and RNS scavenging systems (katG, prxA, hmpA), which were generally thought to be associated with bacterial mutagenesis, we found that deletion of the cyclin gene dps significantly increased the mutation rate (up to 53% at the fifth day post-infection) in V. cholerae. We further determined that the dpsD65A and dpsF46E point mutants showed a similar mutagenesis profile as the Δdps mutant during long-term colonization in mice, which strongly indicated that the antioxidative function of Dps directly contributes to the development of V. cholerae nonmotile mutants. Methionine metabolism pathway may be one of the mechanism for ΔflrA, ΔflrC and ΔrpoN mutant increased colonization in adult mice. Our results revealed a new phenotype in which increased fitness of V. cholerae in the host gut via spontaneous production nonmotile mutants regulated by cyclin Dps, which may represent a novel adaptation strategy for directed evolution of pathogens in the host.
Copyright: © 2023 Luo et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
