

Intermittent antibiotic treatment of bacterial biofilms favors the rapid evolution of resistance
- PMID: 36928386
- PMCID: PMC10020551
- DOI: 10.1038/s42003-023-04601-y
Intermittent antibiotic treatment of bacterial biofilms favors the rapid evolution of resistance
Abstract
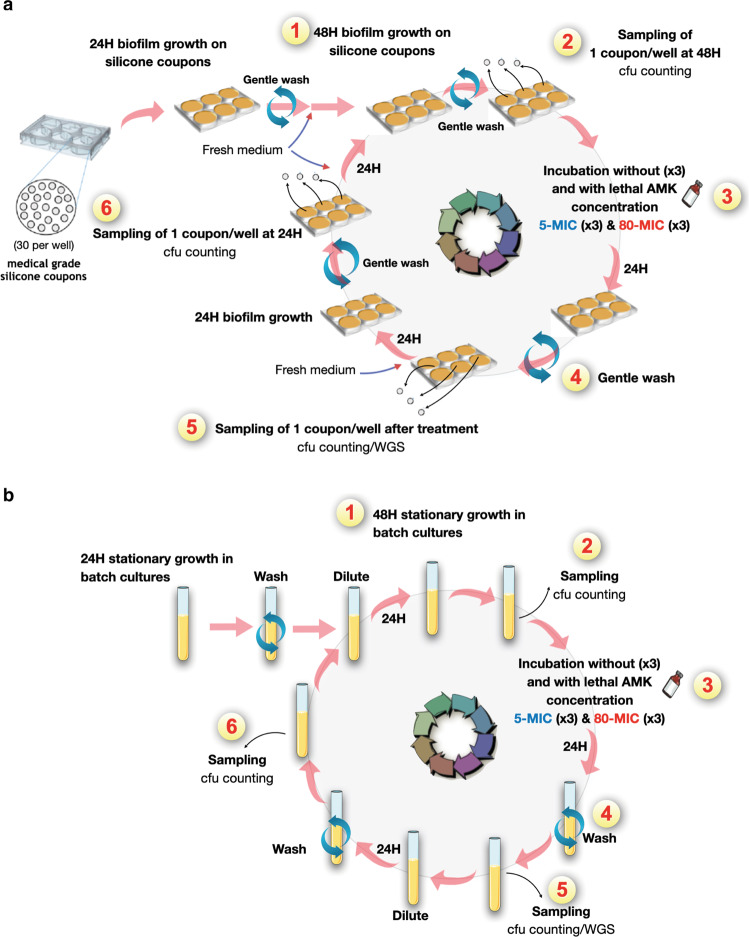
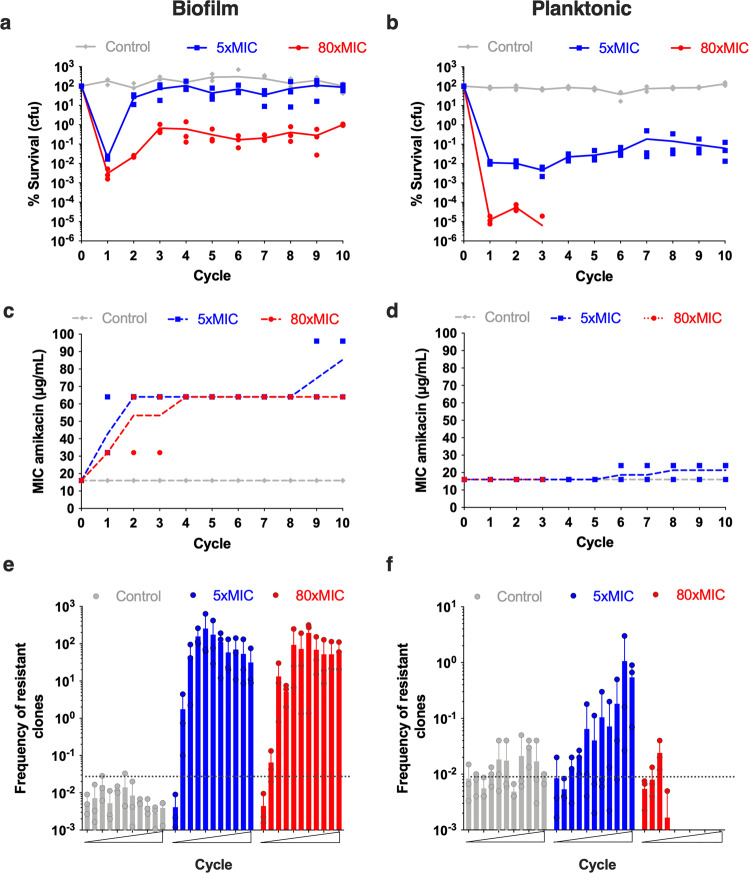
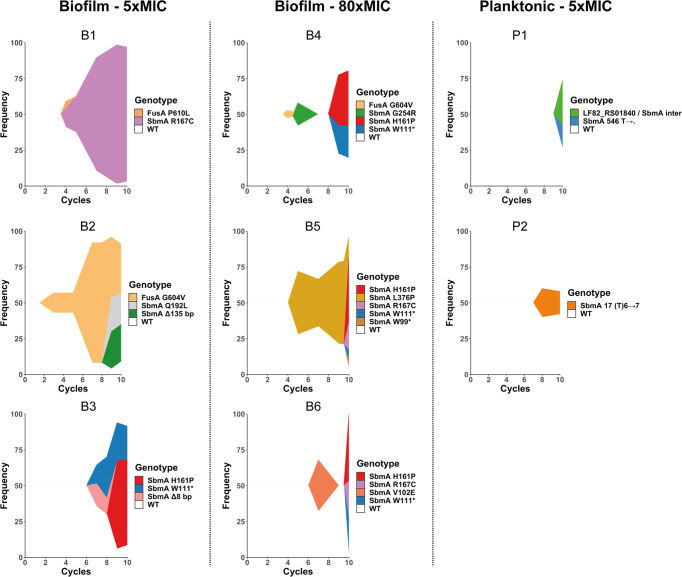
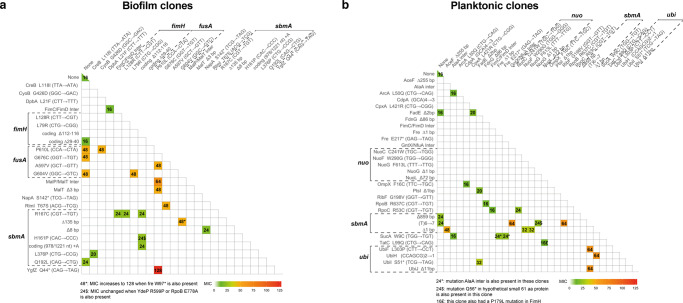
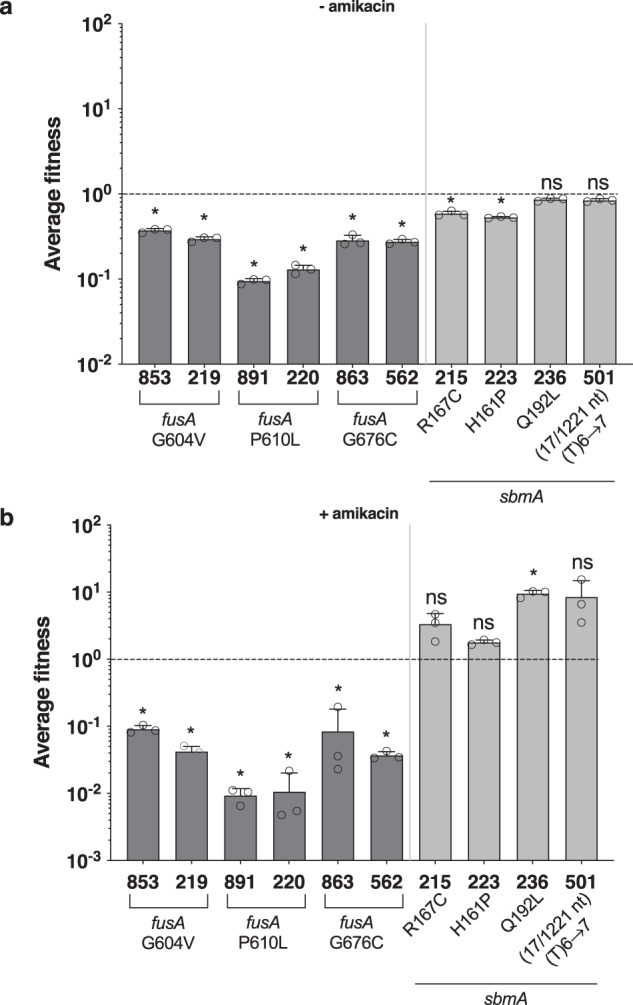
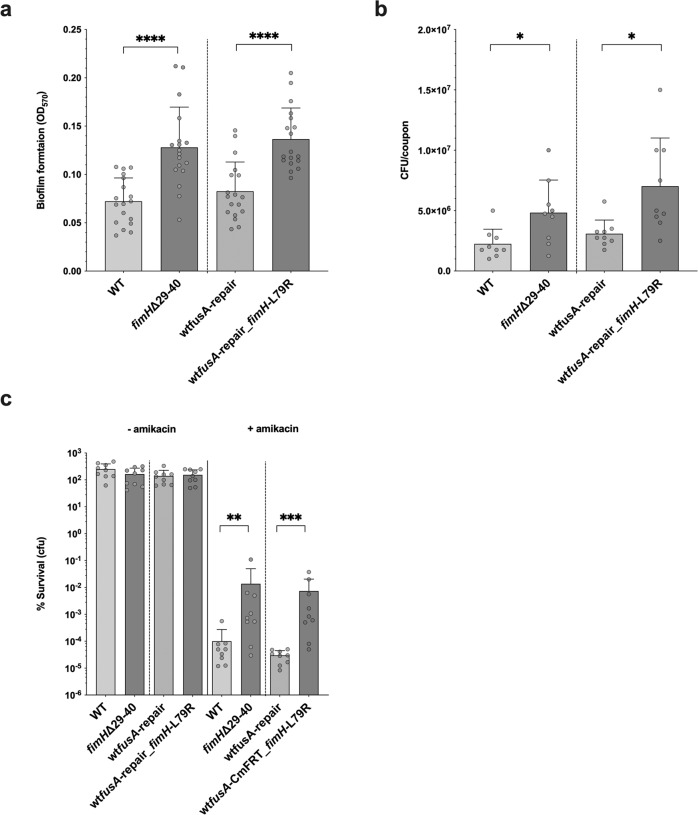
Bacterial antibiotic resistance is a global health concern of increasing importance and intensive study. Although biofilms are a common source of infections in clinical settings, little is known about the development of antibiotic resistance within biofilms. Here, we use experimental evolution to compare selection of resistance mutations in planktonic and biofilm Escherichia coli populations exposed to clinically relevant cycles of lethal treatment with the aminoglycoside amikacin. Consistently, mutations in sbmA, encoding an inner membrane peptide transporter, and fusA, encoding the essential elongation factor G, are rapidly selected in biofilms, but not in planktonic cells. This is due to a combination of enhanced mutation rate, increased adhesion capacity and protective biofilm-associated tolerance. These results show that the biofilm environment favors rapid evolution of resistance and provide new insights into the dynamic evolution of antibiotic resistance in biofilms.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
