

Preclinical models for prediction of immunotherapy outcomes and immune evasion mechanisms in genetically heterogeneous multiple myeloma
- PMID: 36928817
- PMCID: PMC10033443
- DOI: 10.1038/s41591-022-02178-3
Preclinical models for prediction of immunotherapy outcomes and immune evasion mechanisms in genetically heterogeneous multiple myeloma
Abstract
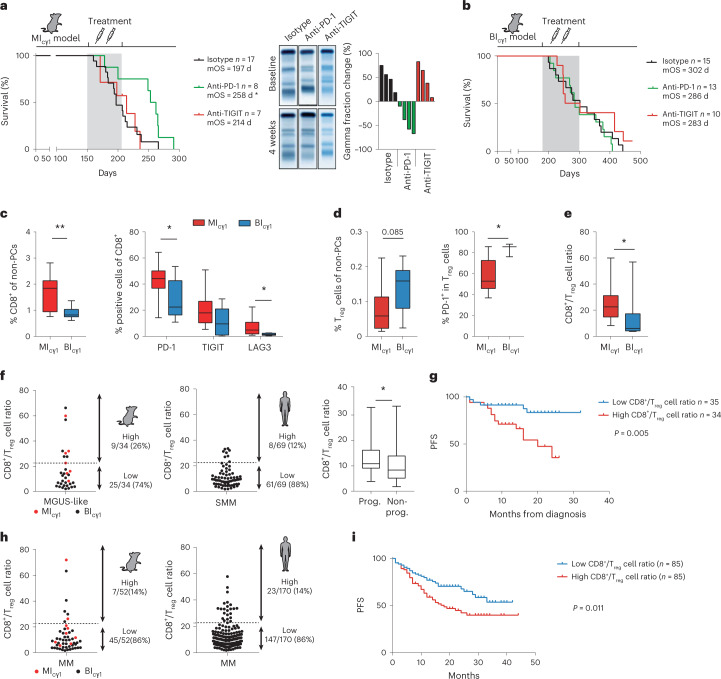
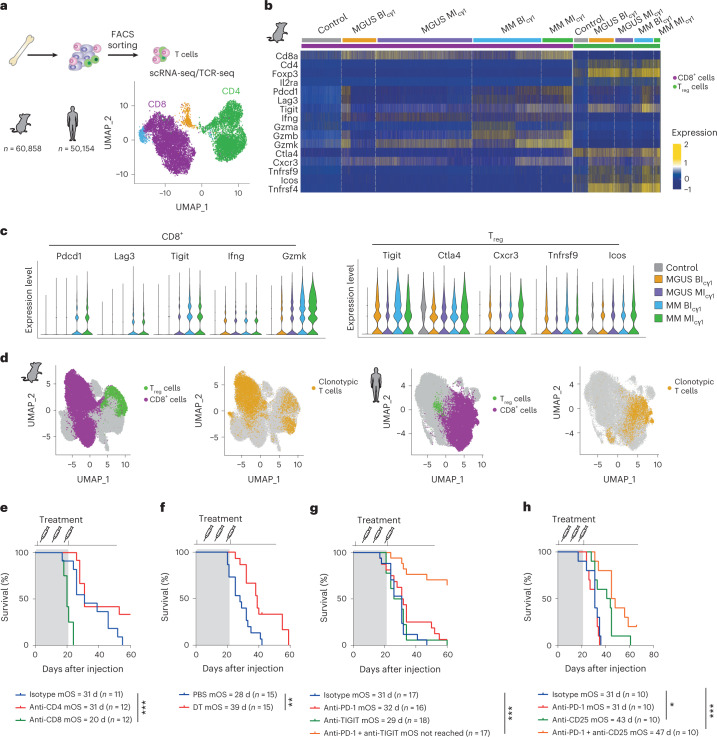
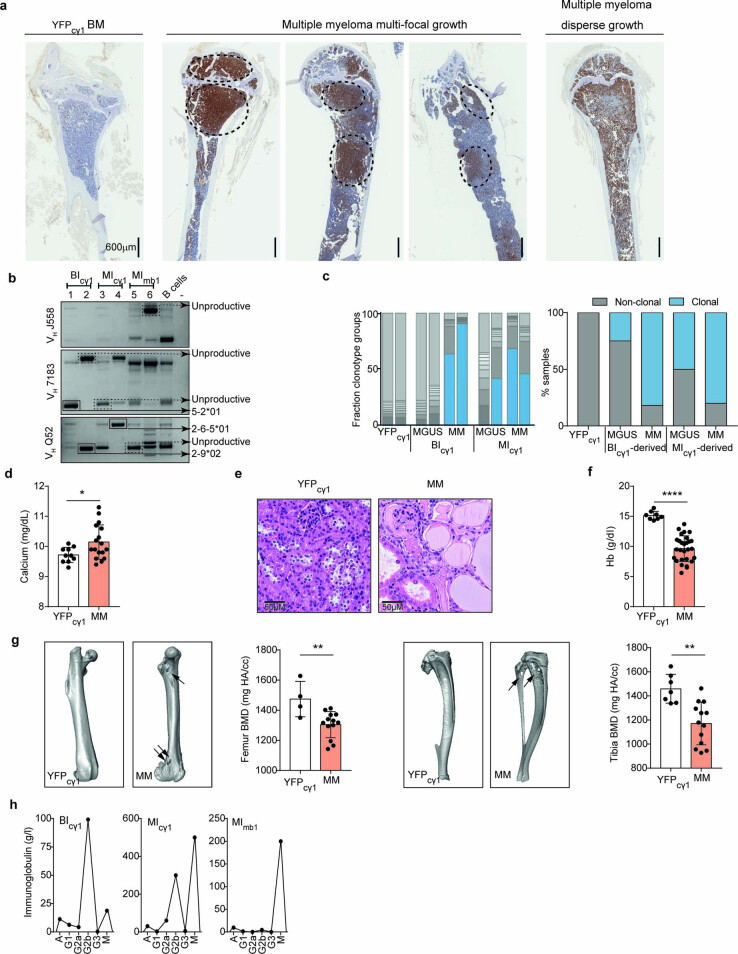
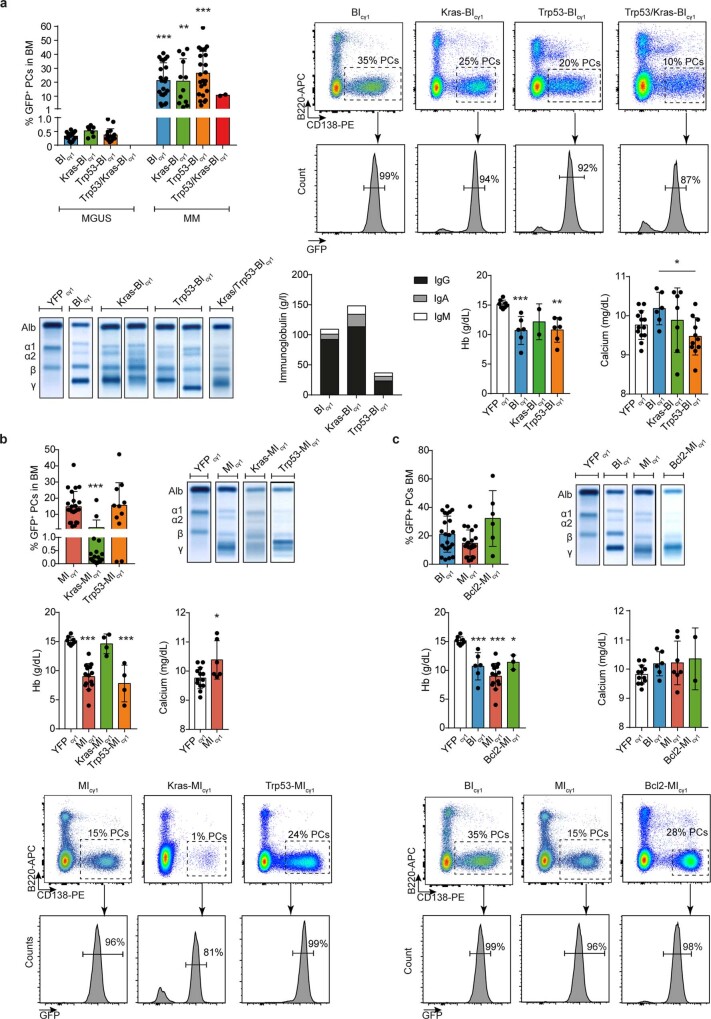
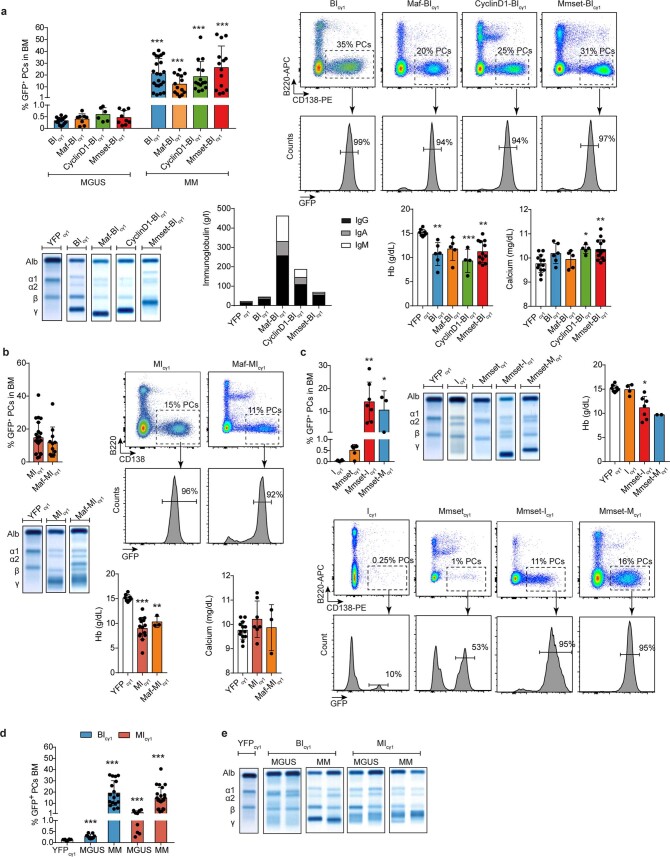
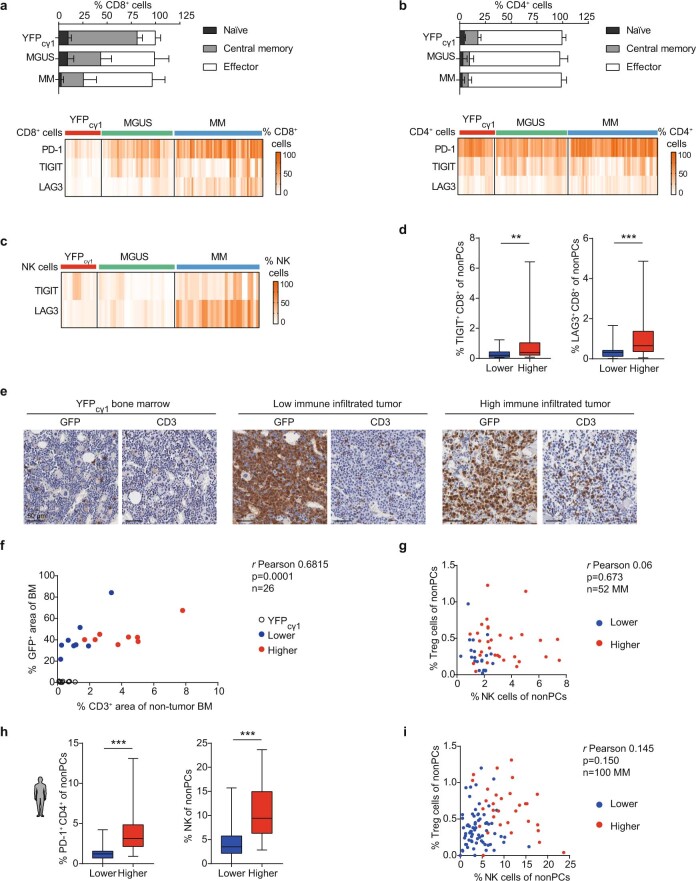
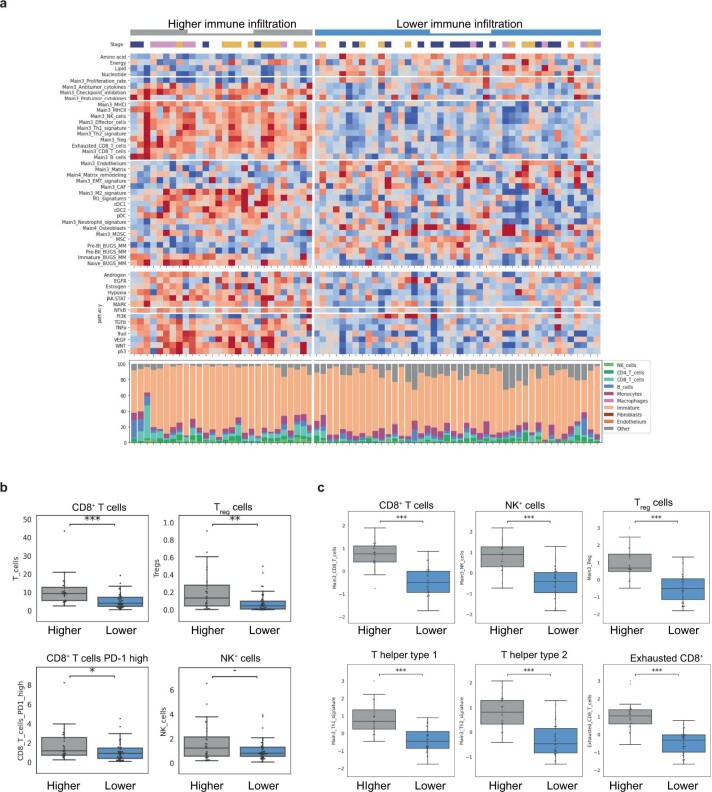
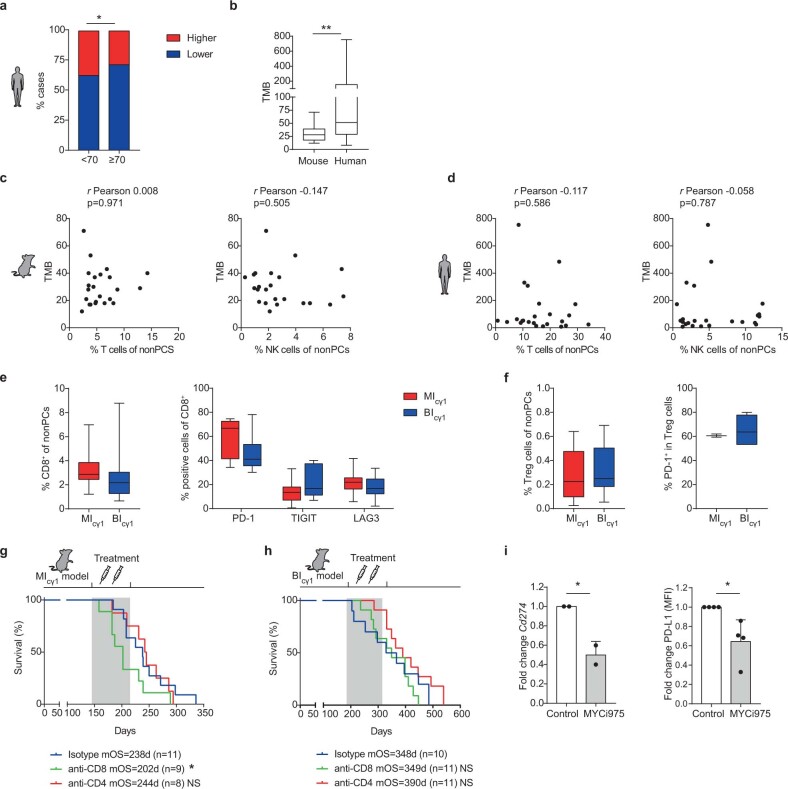
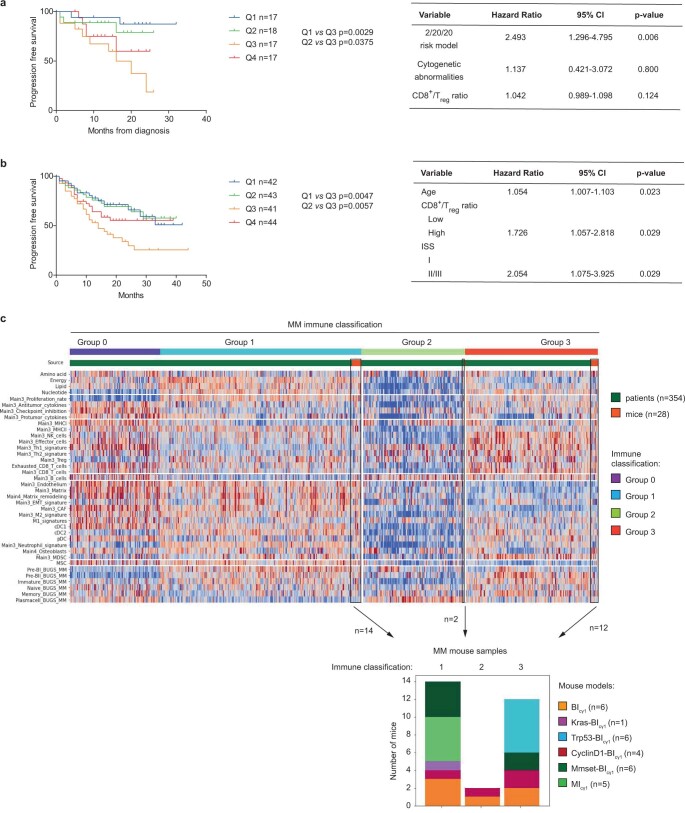
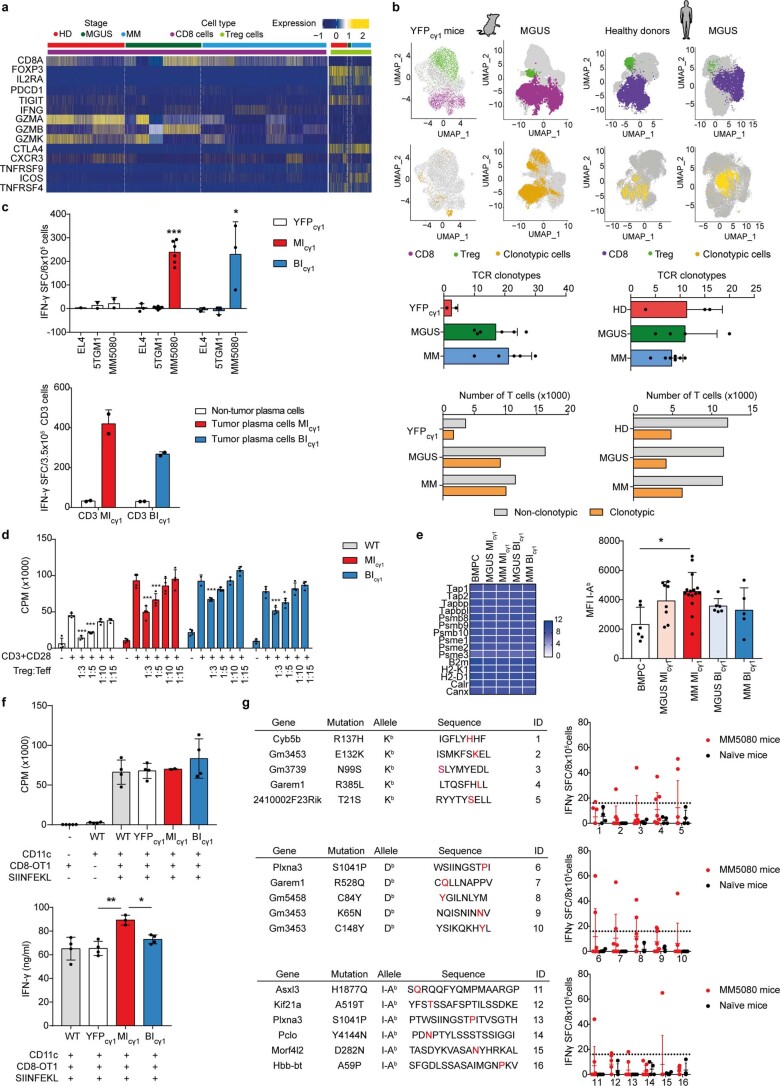
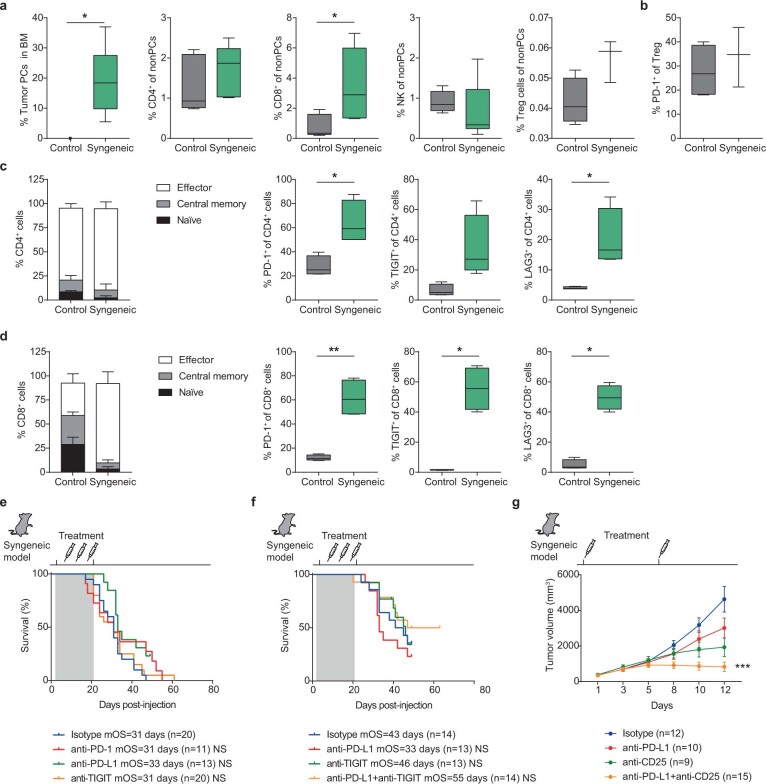
The historical lack of preclinical models reflecting the genetic heterogeneity of multiple myeloma (MM) hampers the advance of therapeutic discoveries. To circumvent this limitation, we screened mice engineered to carry eight MM lesions (NF-κB, KRAS, MYC, TP53, BCL2, cyclin D1, MMSET/NSD2 and c-MAF) combinatorially activated in B lymphocytes following T cell-driven immunization. Fifteen genetically diverse models developed bone marrow (BM) tumors fulfilling MM pathogenesis. Integrative analyses of ∼500 mice and ∼1,000 patients revealed a common MAPK-MYC genetic pathway that accelerated time to progression from precursor states across genetically heterogeneous MM. MYC-dependent time to progression conditioned immune evasion mechanisms that remodeled the BM microenvironment differently. Rapid MYC-driven progressors exhibited a high number of activated/exhausted CD8+ T cells with reduced immunosuppressive regulatory T (Treg) cells, while late MYC acquisition in slow progressors was associated with lower CD8+ T cell infiltration and more abundant Treg cells. Single-cell transcriptomics and functional assays defined a high ratio of CD8+ T cells versus Treg cells as a predictor of response to immune checkpoint blockade (ICB). In clinical series, high CD8+ T/Treg cell ratios underlie early progression in untreated smoldering MM, and correlated with early relapse in newly diagnosed patients with MM under Len/Dex therapy. In ICB-refractory MM models, increasing CD8+ T cell cytotoxicity or depleting Treg cells reversed immunotherapy resistance and yielded prolonged MM control. Our experimental models enable the correlation of MM genetic and immunological traits with preclinical therapy responses, which may inform the next-generation immunotherapy trials.
© 2023. The Author(s).
Conflict of interest statement
M.C. has received honoraria from participation in advisory boards from Oncopeptides, Novartis, Janssen and Pfizer; and has licensed intellectual property through Mayo Clinical Ventures 2013-058 ‘Genetically engineered mouse model of MM without LoxP sites (Vk*MycCwoLoxP) and transplantable cell lines’. L.B. has received honoraria from participation in advisory boards from Oncopeptides, Novartis, Janssen and Pfizer; and has licensed intellectual property through Mayo Clinical Ventures 2013-058 ‘Genetically engineered mouse model of MM without LoxP sites (Vk*MycCwoLoxP) and transplantable cell lines’. P.R.-O. has received honoraria from advisory boards from Pfizer, BMS, Janssen, GSK, Kite, Sanofi and Oncopeptides; and honoraria from lectures from GSK, Janssen, BMS, Regeneron, Amgen and Oncopeptides. S.M.R. is an employee of Roche/Genentech. E.A.L. is an employee of Roche/Genentech. M.A. is an employee of Roche, and has patent applications on CD25 monoclonal antibody with relevance to this work: WO/2018/167104 and US20190284287 (filed by Cancer Research Technology Limited and Tusk Therapeutics). M.A. has shares in the companies to which the patent belongs. O.K. is an employee of BostonGene. J.S.M. is a consultant and an advisory board member for (on behalf of his Institution) Amgen, BMS, Celgene, Haemalogix, Janssen, MSD, Novartis, Takeda, Sanofi, Roche, Abbvie, GlaxoSmithKline, Regeneron, SecuraBio and Karyopharm. B.P. reports honoraria for lectures from and membership on advisory boards with Adaptive, Amgen, Becton Dickinson, Bristol Myers Squibb-Celgene, Janssen, Merck, Novartis, Roche, Sanofi and Takeda; unrestricted grants from Bristol Myers Squibb-Celgene, EngMab, Roche, Sanofi and Takeda; and consultancy for Bristol Myers Squibb-Celgene, Janssen, Sanofi and Takeda. J.A.M.-C. reports funding for research from Roche/Genentech, Bristol Myers Squibb-Celgene, Janssen, Priothera and Palleon. A patent on the generation and use of the mouse MM models as immunotherapy platforms, entitled ‘Genetically engineered animal models for multiple myeloma’ (application no. EP22382736.1) was filed on 27 July 2022. The remaining authors declare no competing interests.
Figures
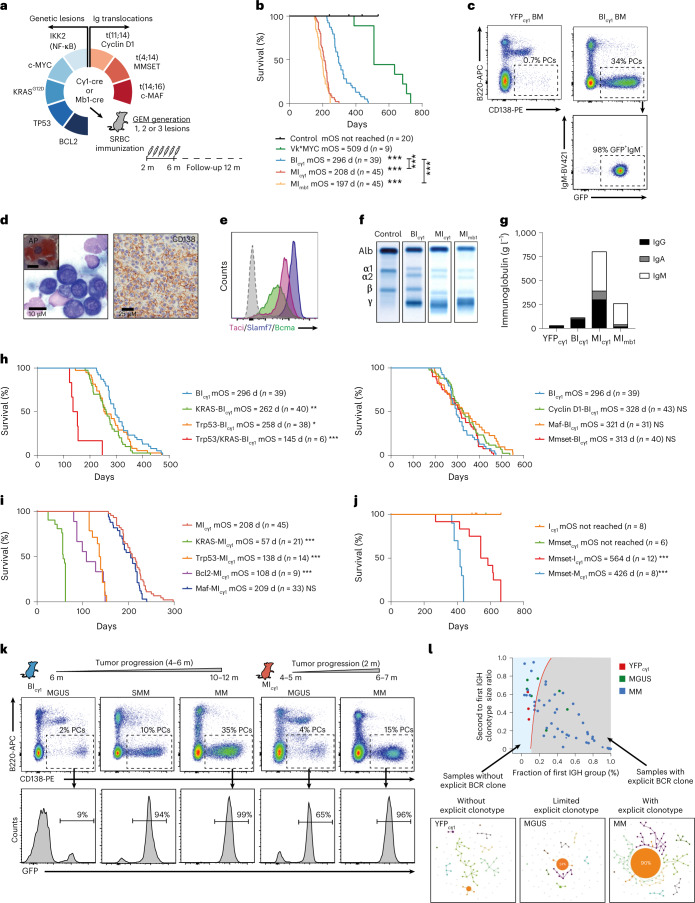
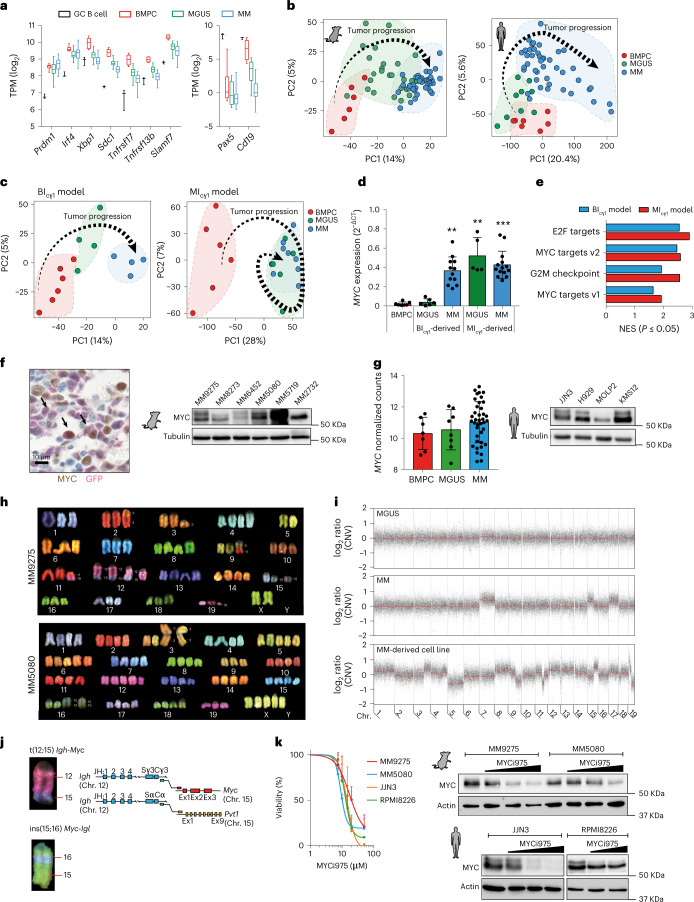
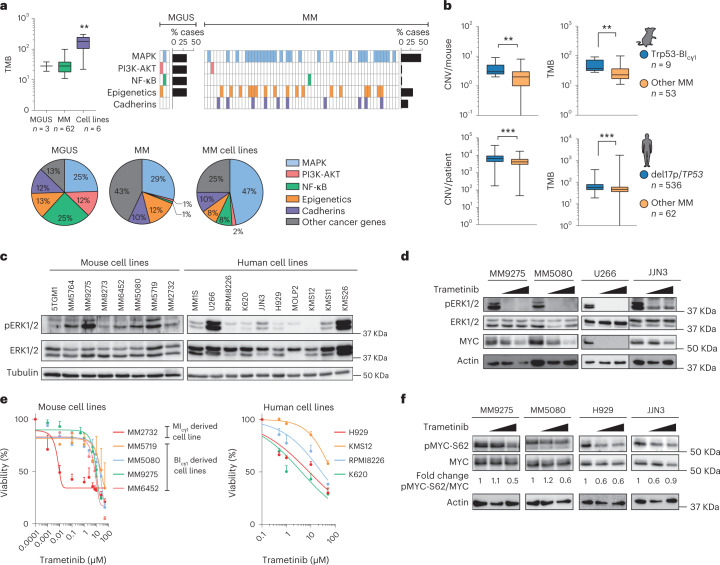
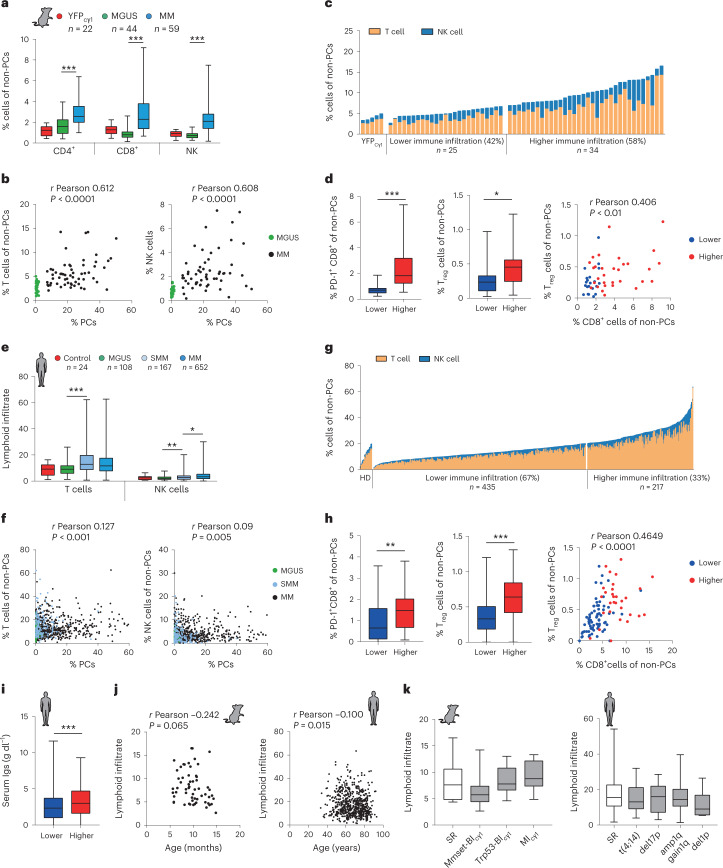
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
