

Evolutionary Trajectories are Contingent on Mitonuclear Interactions
- PMID: 36929911
- PMCID: PMC10072823
- DOI: 10.1093/molbev/msad061
Evolutionary Trajectories are Contingent on Mitonuclear Interactions
Abstract
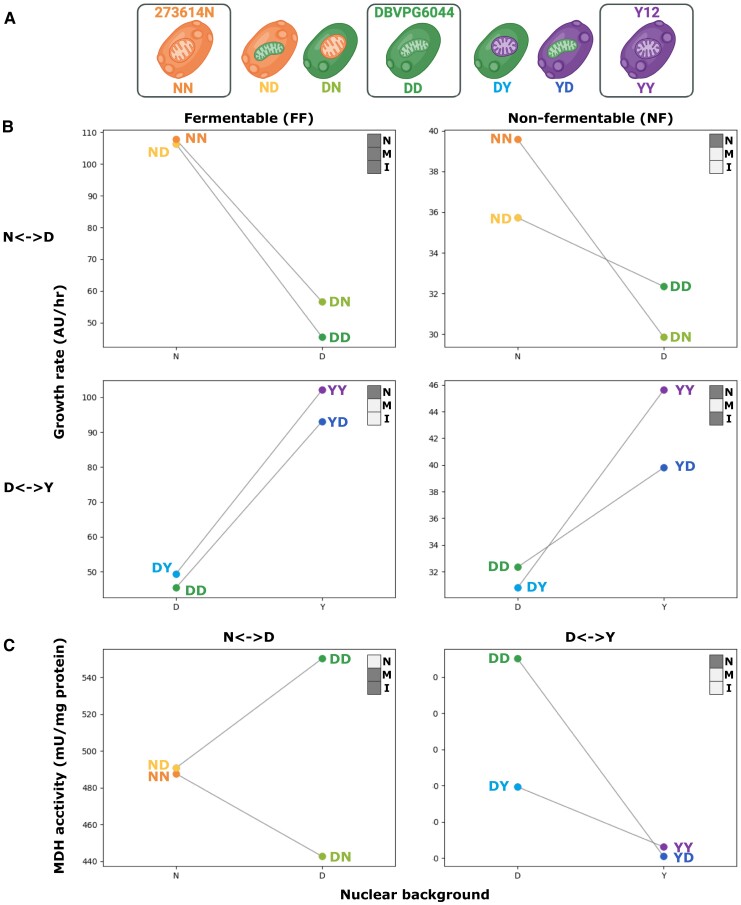
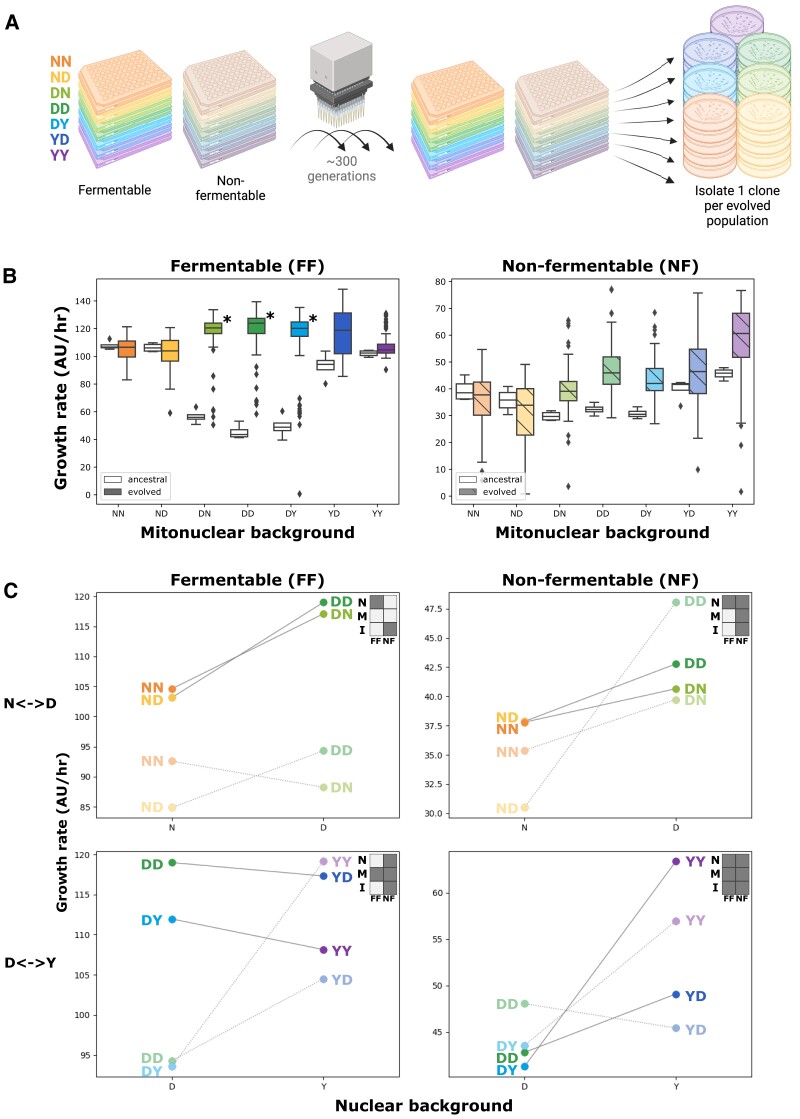
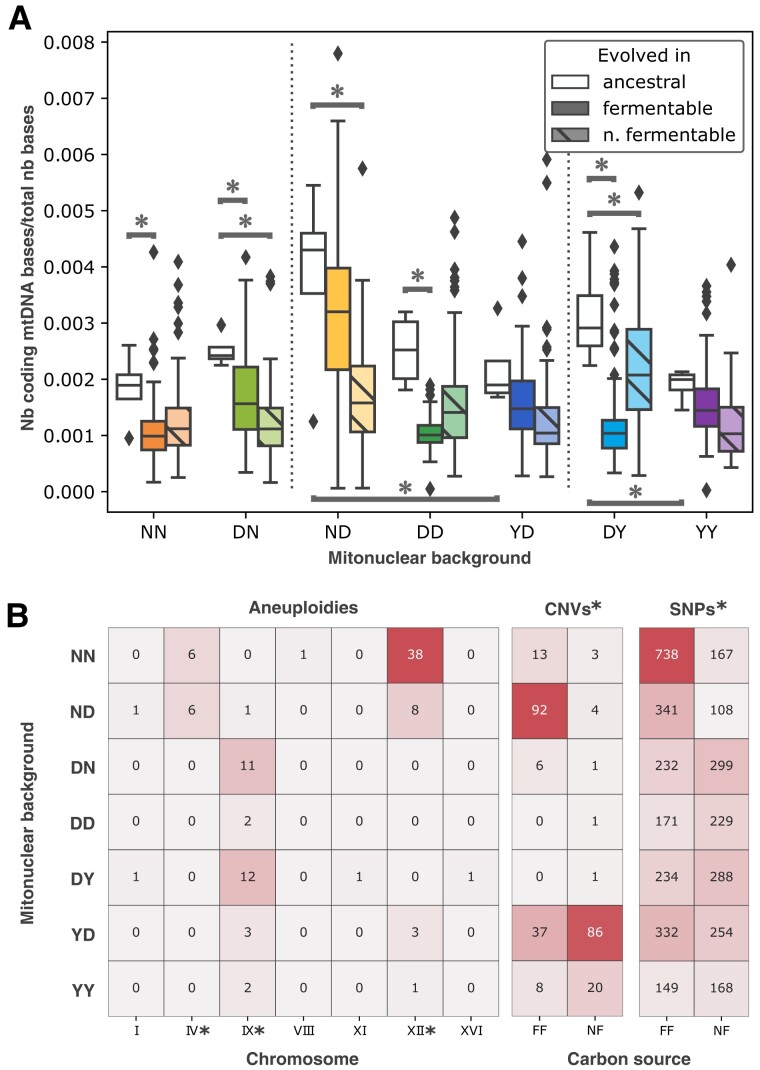
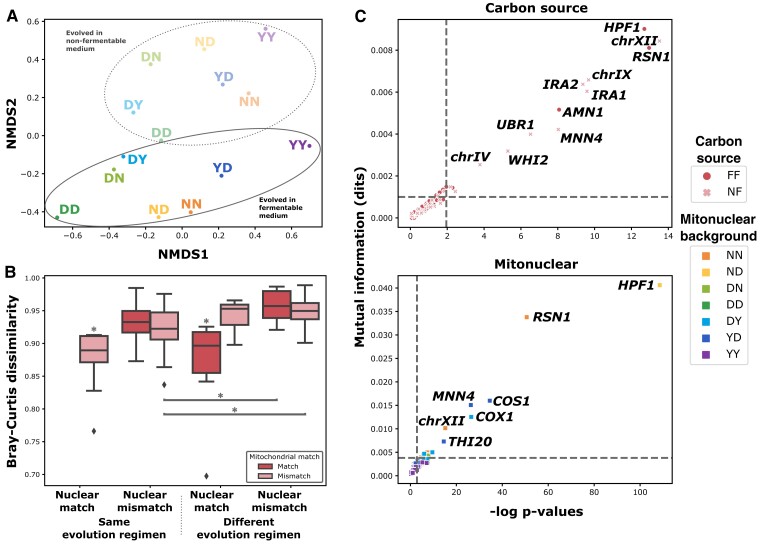
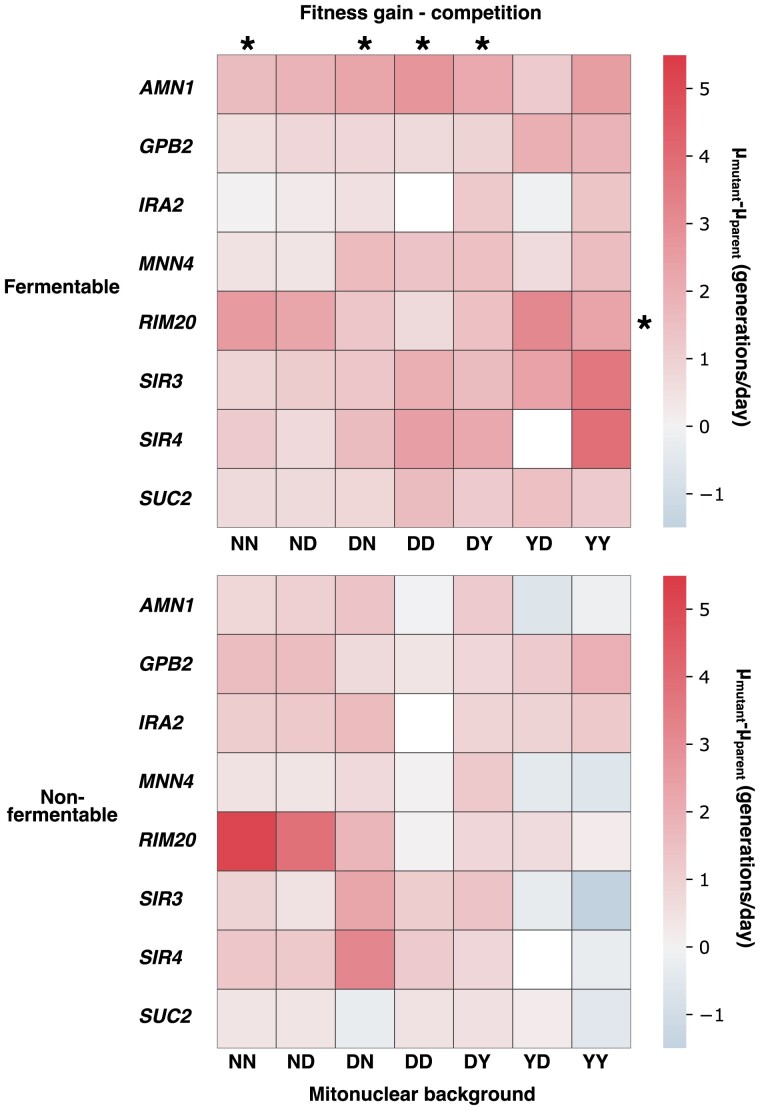
Critical mitochondrial functions, including cellular respiration, rely on frequently interacting components expressed from both the mitochondrial and nuclear genomes. The fitness of eukaryotic organisms depends on a tight collaboration between both genomes. In the face of an elevated rate of evolution in mtDNA, current models predict that the maintenance of mitonuclear compatibility relies on compensatory evolution of the nuclear genome. Mitonuclear interactions would therefore exert an influence on evolutionary trajectories. One prediction from this model is that the same nuclear genome evolving with different mitochondrial haplotypes would follow distinct molecular paths toward higher fitness. To test this prediction, we submitted 1,344 populations derived from 7 mitonuclear genotypes of Saccharomyces cerevisiae to >300 generations of experimental evolution in conditions that either select for a mitochondrial function or do not strictly require respiration for survival. Performing high-throughput phenotyping and whole-genome sequencing on independently evolved individuals, we identified numerous examples of gene-level evolutionary convergence among populations with the same mitonuclear background. Phenotypic and genotypic data on strains derived from this evolution experiment identify the nuclear genome and the environment as the main determinants of evolutionary divergence, but also show a modulating role for the mitochondrial genome exerted both directly and via interactions with the two other components. We finally recapitulated a subset of prominent loss-of-function alleles in the ancestral backgrounds and confirmed a generalized pattern of mitonuclear-specific and highly epistatic fitness effects. Together, these results demonstrate how mitonuclear interactions can dictate evolutionary divergence of populations with identical starting nuclear genotypes.
Keywords: evolutionary convergence; experimental evolution; mitochondria; mitonuclear evolution.
© The Author(s) 2023. Published by Oxford University Press on behalf of Society for Molecular Biology and Evolution.
Figures
References
-
- Andrews S. 2010. FastQC: a quality control tool for high throughput sequence data. Available from:http://www.bioinformatics.babraham.ac.uk/projects/fastqc/
-
- Barreto FS, Watson ET, Lima TG, Willett CS, Edmands S, Li W, Burton RS. 2018. Genomic signatures of mitonuclear coevolution across populations of Tigriopus californicus. Nat Ecol Evol. 2:1250–1257. - PubMed
-
- Barrientos A, Kenyon L, Moraes CT. 1998. Human xenomitochondrial cybrids. J Biol Chem. 273:14210–14217. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
