

The η-secretase-derived APP fragment ηCTF is localized in Golgi, endosomes and extracellular vesicles and contributes to Aβ production
- PMID: 36930302
- PMCID: PMC10023608
- DOI: 10.1007/s00018-023-04737-4
The η-secretase-derived APP fragment ηCTF is localized in Golgi, endosomes and extracellular vesicles and contributes to Aβ production
Abstract
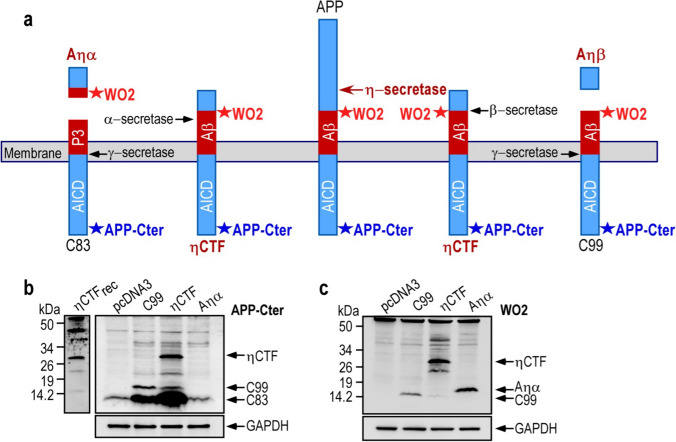
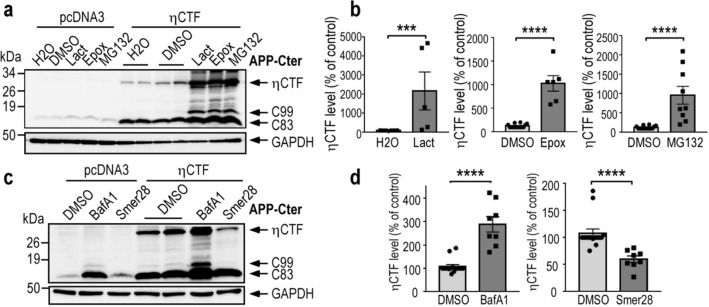
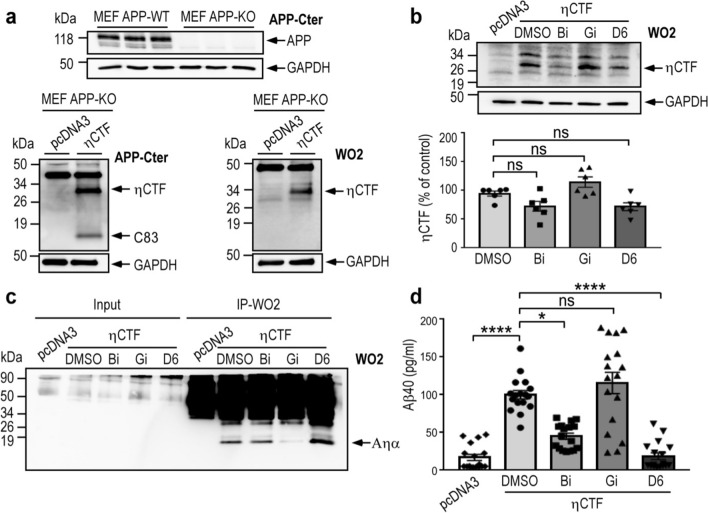
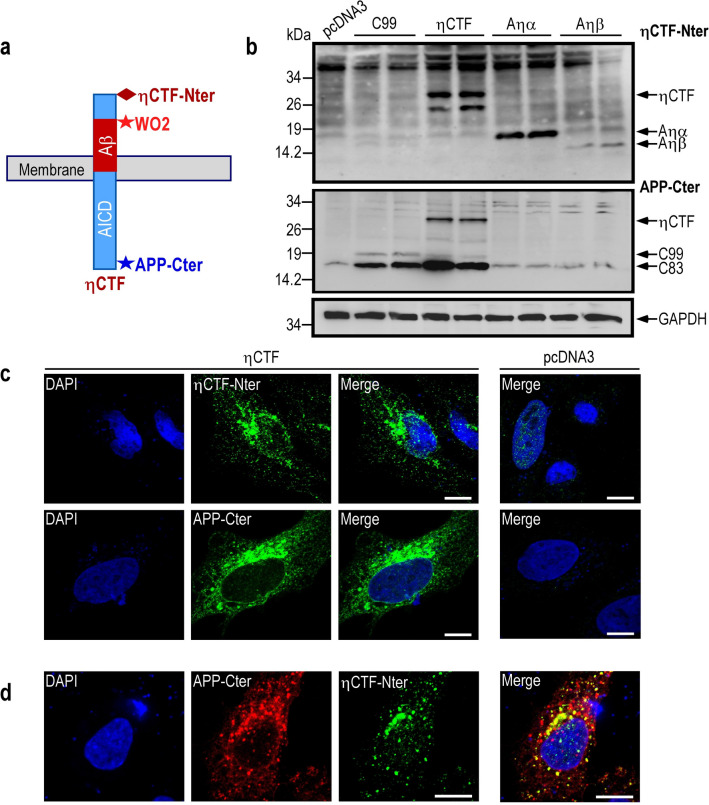
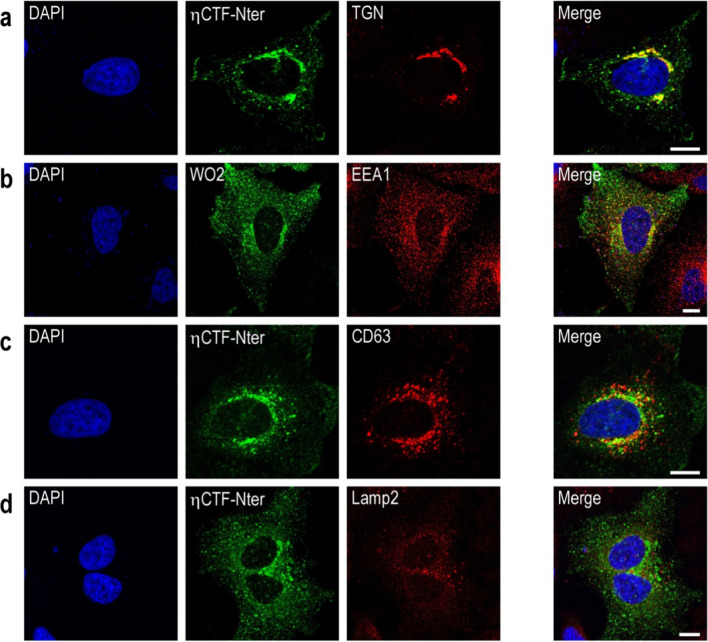
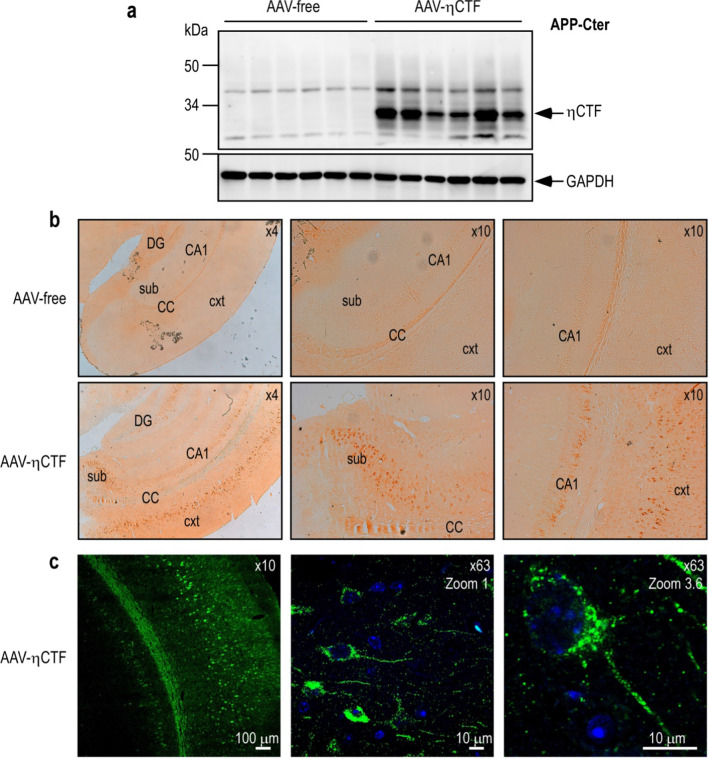
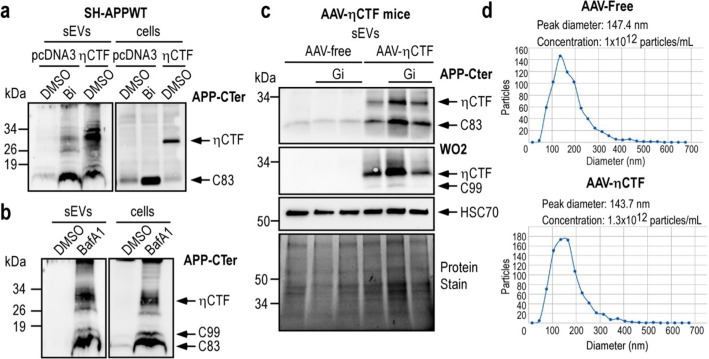
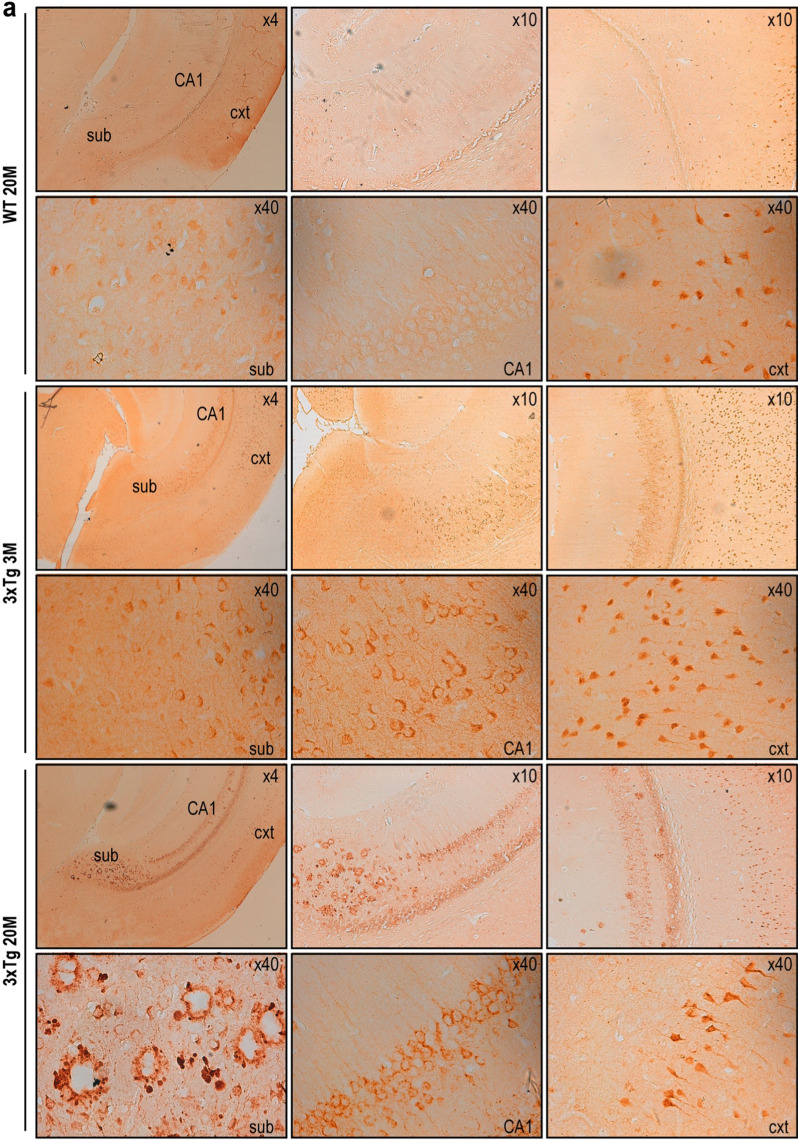
The processing of the amyloid precursor protein (APP) is one of the key events contributing to Alzheimer's disease (AD) etiology. Canonical cleavages by β- and γ-secretases lead to Aβ production which accumulate in amyloid plaques. Recently, the matrix metalloprotease MT5-MMP, referred to as η-secretase, has been identified as a novel APP cleaving enzyme producing a transmembrane fragment, ηCTF that undergoes subsequent cleavages by α- and β-secretases yielding the Aηα and Aηβ peptides, respectively. The functions and contributions of ηCTF and its related fragments to AD pathology are poorly understood. In this study, we designed a novel immunological probe referred to as ηCTF-NTer antibody that specifically interacts with the N-terminal part of ηCTF targeting ηCTF, Aηα, Aηβ but not C99, C83 and Aβ. We examined the fate and localization of ηCTF fragment in various cell models and in mice. We found that overexpressed ηCTF undergoes degradation in the proteasomal and autophagic pathways and accumulates mainly in the Golgi and in endosomes. Moreover, we observed the presence of ηCTF in small extracellular vesicles purified from neuroblastoma cells or from mouse brains expressing ηCTF. Importantly, the expression of ηCTF in fibroblasts devoid on APP leads to Aβ production demonstrating its contribution to the amyloidogenic pathway. Finally, we observed an ηCTF-like immunoreactivity around amyloid plaques and an age-dependent accumulation of ηCTF in the triple-transgenic mouse AD model. Thus, our study suggests that the ηCTF fragment likely contributes to AD pathology by its exosomal spreading and involvement in Aβ production.
Keywords: Alzheimer’s disease; Autophagic degradation; Aβ-production; Endosomes; Extracellular vesicles; ηCTF.
© 2023. The Author(s).
Conflict of interest statement
The authors have nothing to disclose and declare no financial conflict of interest.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
