

Downregulation of Hsp90 and the antimicrobial peptide Mtk suppresses poly(GR)-induced neurotoxicity in C9ORF72-ALS/FTD
- PMID: 36931278
- PMCID: PMC10264157
- DOI: 10.1016/j.neuron.2023.02.029
Downregulation of Hsp90 and the antimicrobial peptide Mtk suppresses poly(GR)-induced neurotoxicity in C9ORF72-ALS/FTD
Abstract
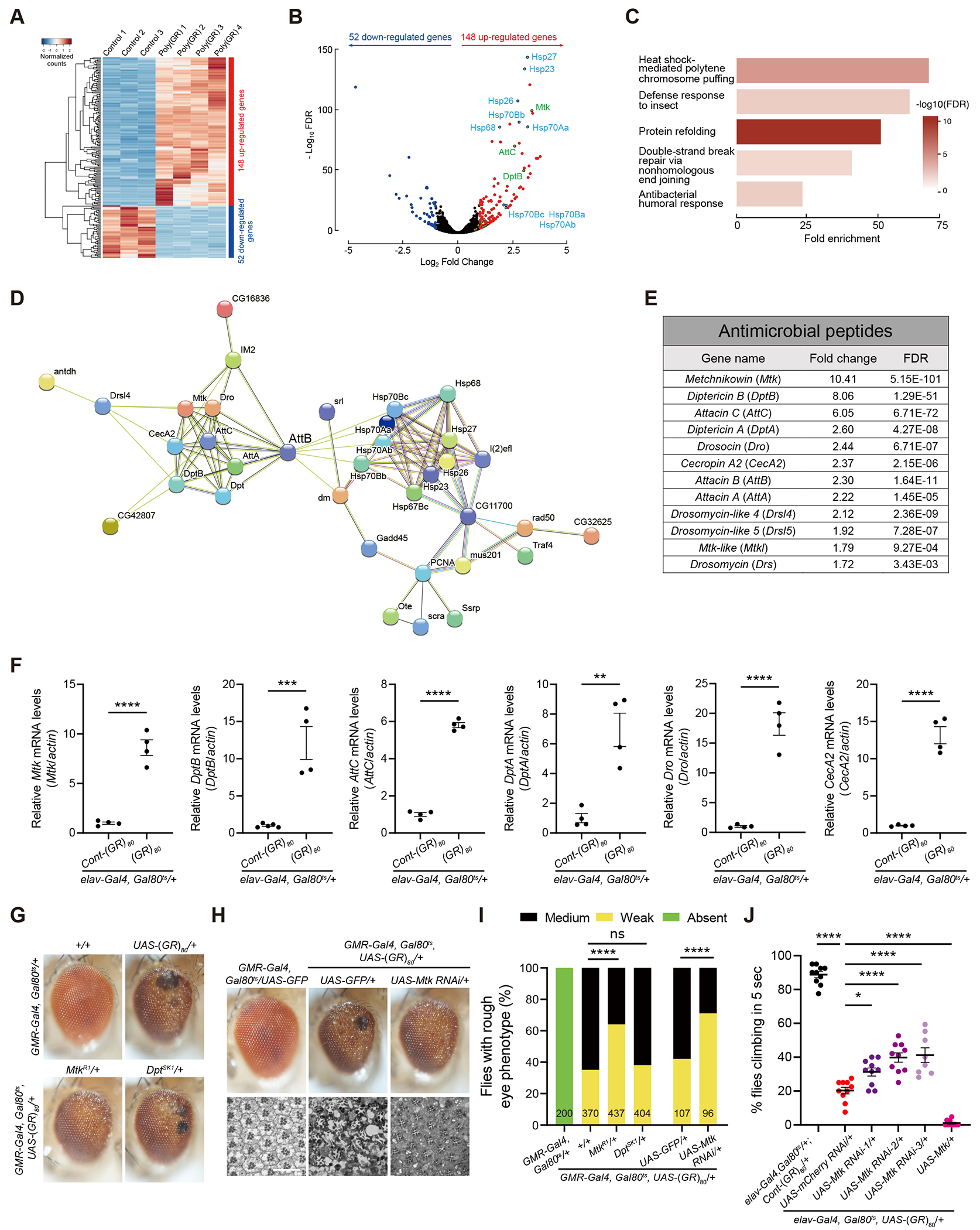
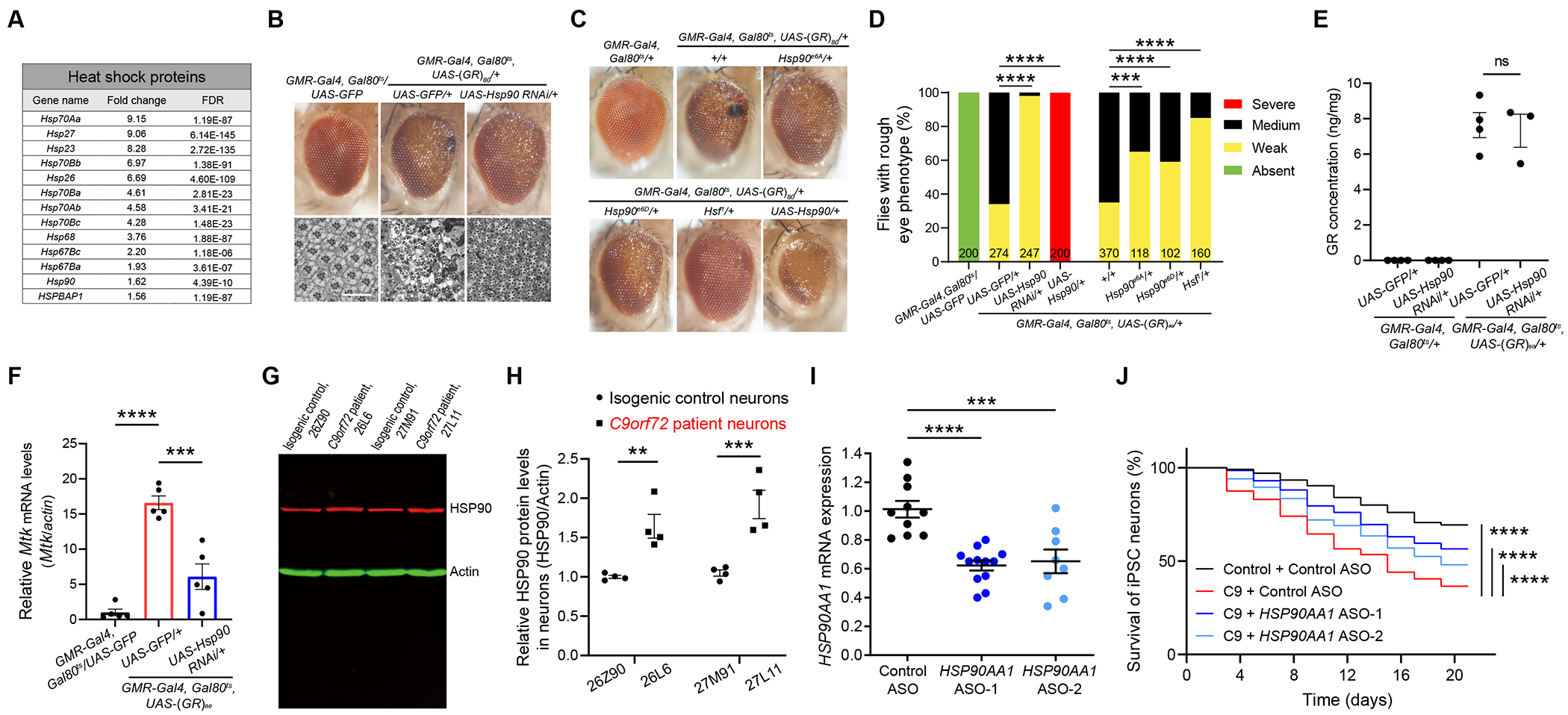
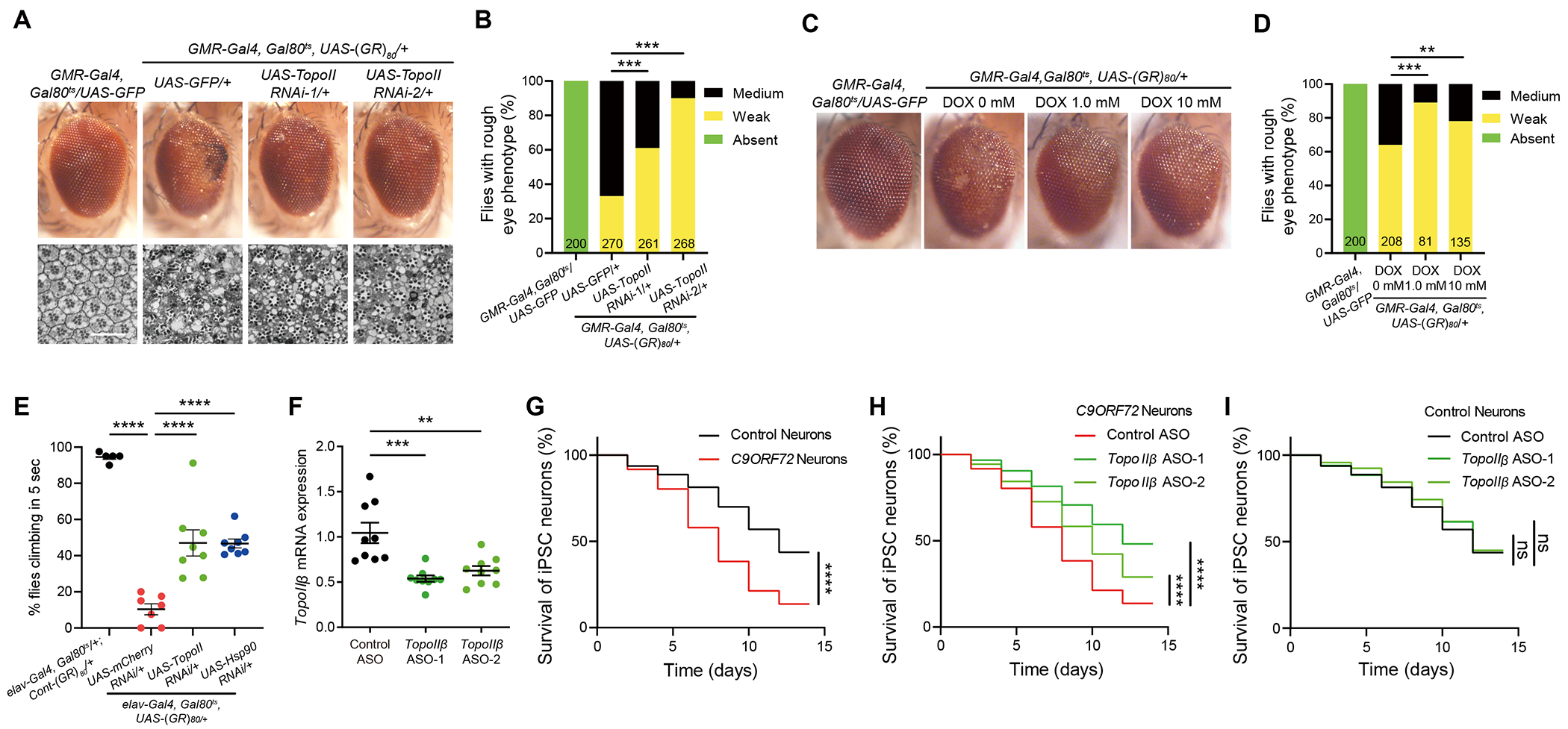
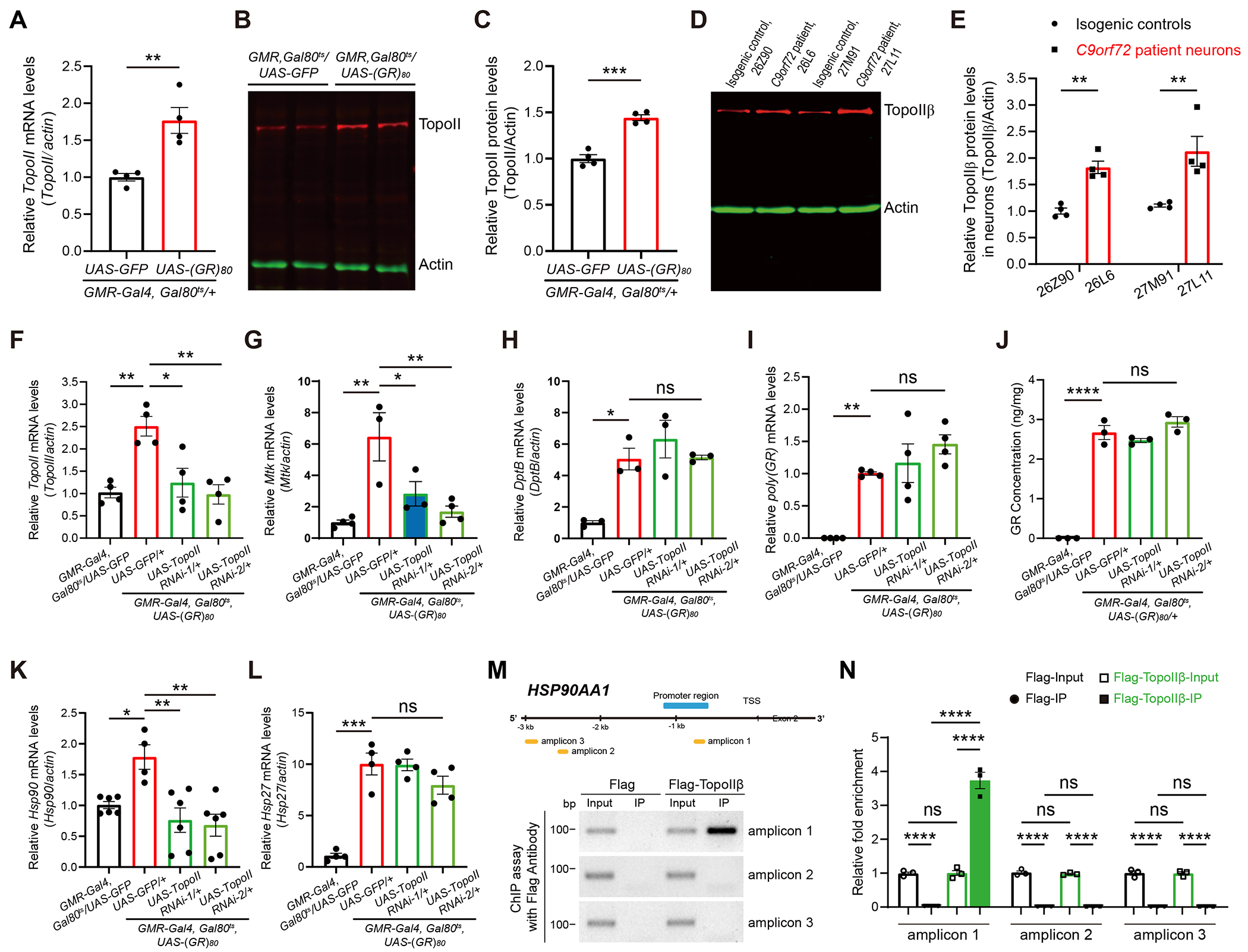
GGGGCC repeat expansion in the C9ORF72 gene is the most common genetic cause of amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD). Repeat RNAs can be translated into dipeptide repeat proteins, including poly(GR), whose mechanisms of action remain largely unknown. In an RNA-seq analysis of poly(GR) toxicity in Drosophila, we found that several antimicrobial peptide genes, such as metchnikowin (Mtk), and heat shock protein (Hsp) genes are activated. Mtk knockdown in the fly eye or in all neurons suppresses poly(GR) neurotoxicity. These findings suggest a cell-autonomous role of Mtk in neurodegeneration. Hsp90 knockdown partially rescues both poly(GR) toxicity in flies and neurodegeneration in C9ORF72 motor neurons derived from induced pluripotent stem cells (iPSCs). Topoisomerase II (TopoII) regulates poly(GR)-induced upregulation of Hsp90 and Mtk. TopoII knockdown also suppresses poly(GR) toxicity in Drosophila and improves survival of C9ORF72 iPSC-derived motor neurons. These results suggest potential novel therapeutic targets for C9ORF72-ALS/FTD.
Keywords: ALS; C9ORF72; DPR; FTD; Hsp90; Mtk; TopoII; antimicrobial peptide; neurodegeneration; poly(GR).
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests F.-B.G. has an active research agreement with, and receives funding from, Stealth BioTherapeutics. Z.W. co-founded Rgenta Therapeutics, and she serves as a scientific advisor for the company and is a member of its board. J.I. is a co-founder of AcuraStem, Inc. and Modulo Bio, serves on the scientific advisory boards of AcuraStem, Spinogenix, Synapticure, and Vesalius Therapeutics, and is employed at BioMarin Pharmaceutical.
Figures
References
-
- DeJesus-Hernandez M, Mackenzie IR, Boeve BF, Boxer AL, Baker M, Rutherford NJ, Nicholson AM, Finch NA, Flynn H, Adamson J, et al. (2011). Expanded GGGGCC hexanucleotide repeat in noncoding region of C9ORF72 causes chromosome 9p-linked FTD and ALS. Neuron 72, 245–256. 10.1016/j.neuron.2011.09.011. - DOI - PMC - PubMed
-
- Renton AE, Majounie E, Waite A, Simón-Sánchez J, Rollinson S, Gibbs JR, Schymick JC, Laaksovirta H, van Swieten JC, Myllykangas L, et al. (2011). A hexanucleotide repeat expansion in C9ORF72 is the cause of chromosome 9p21-linked ALS-FTD. Neuron 72, 257–268. 10.1016/j.neuron.2011.09.010. - DOI - PMC - PubMed
-
- Ash PE, Bieniek KF, Gendron TF, Caulfield T, Lin WL, Dejesus-Hernandez M, van Blitterswijk MM, Jansen-West K, Paul JW 3rd, Rademakers R, et al. (2013). Unconventional translation of C9ORF72 GGGGCC expansion generates insoluble polypeptides specific to c9FTD/ALS. Neuron 77, 639–646. 10.1016/j.neuron.2013.02.004. - DOI - PMC - PubMed
-
- Zu T, Liu Y, Bañez-Coronel M, Reid T, Pletnikova O, Lewis J, Miller TM, Harms MB, Falchook AE, Subramony SH, et al. (2013). RAN proteins and RNA foci from antisense transcripts in C9ORF72 ALS and frontotemporal dementia. Proc. Natl. Acad. Sci. USA 110, 4968–4977. 10.1073/pnas.1315438110. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
