

Molecular basis of Q-length selectivity for the MW1 antibody-huntingtin interaction
- PMID: 36931390
- PMCID: PMC10124945
- DOI: 10.1016/j.jbc.2023.104616
Molecular basis of Q-length selectivity for the MW1 antibody-huntingtin interaction
Abstract
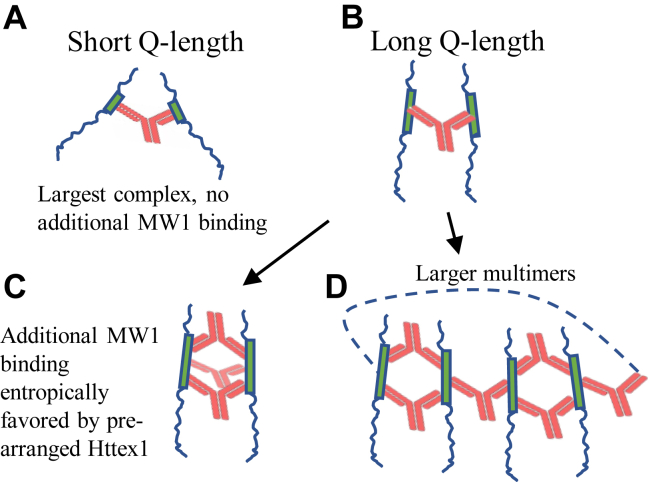
Huntington's disease is caused by a polyglutamine (polyQ) expansion in the huntingtin protein. Huntingtin exon 1 (Httex1), as well as other naturally occurring N-terminal huntingtin fragments with expanded polyQ are prone to aggregation, forming potentially cytotoxic oligomers and fibrils. Antibodies and other N-terminal huntingtin binders are widely explored as biomarkers and possible aggregation-inhibiting therapeutics. A monoclonal antibody, MW1, is known to preferentially bind to huntingtin fragments with expanded polyQ lengths, but the molecular basis of the polyQ length specificity remains poorly understood. Using solution NMR, electron paramagnetic resonance, and other biophysical methods, we investigated the structural features of the Httex1-MW1 interaction. Rather than recognizing residual α-helical structure, which is promoted by expanded Q-lengths, MW1 caused the formation of a new, non-native, conformation in which the entire polyQ is largely extended. This non-native polyQ structure allowed the formation of large mixed Httex1-MW1 multimers (600-2900 kD), when Httex1 with pathogenic Q-length (Q46) was used. We propose that these multivalent, entropically favored interactions, are available only to proteins with longer Q-lengths and represent a major factor governing the Q-length preference of MW1. The present study reveals that it is possible to target proteins with longer Q-lengths without having to stabilize a natively favored conformation. Such mechanisms could be exploited in the design of other Q-length specific binders.
Keywords: Huntington’s disease; MW1; antibody; biomarkers; electron paramagnetic resonance; polyglutamine; protein aggregation; protein conformation; protein-aggregation inhibitors.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare no conflict of interest with the contents of this article.
Figures






References
-
- Walker F.O. Huntington's disease. Lancet. 2007;369:218–228. - PubMed
-
- Hoffner G., Djian P. Polyglutamine aggregation in huntington disease: does structure determine toxicity? Mol. Neurobiol. 2015;52:1297–1314. - PubMed
-
- Mangiarini L., Sathasivam K., Seller M., Cozens B., Harper A., Hetherington C., et al. Exon 1 of the HD gene with an expanded CAG repeat is sufficient to cause a progressive neurological phenotype in transgenic mice. Cell. 1996;87:493–506. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
