

Lineages to circuits: the developmental and evolutionary architecture of information channels into the central complex
- PMID: 36932234
- PMCID: PMC10354165
- DOI: 10.1007/s00359-023-01616-y
Lineages to circuits: the developmental and evolutionary architecture of information channels into the central complex
Abstract
The representation and integration of internal and external cues is crucial for any organism to execute appropriate behaviors. In insects, a highly conserved region of the brain, the central complex (CX), functions in the representation of spatial information and behavioral states, as well as the transformation of this information into desired navigational commands. How does this relatively invariant structure enable the incorporation of information from the diversity of anatomical, behavioral, and ecological niches occupied by insects? Here, we examine the input channels to the CX in the context of their development and evolution. Insect brains develop from ~ 100 neuroblasts per hemisphere that divide systematically to form "lineages" of sister neurons, that project to their target neuropils along anatomically characteristic tracts. Overlaying this developmental tract information onto the recently generated Drosophila "hemibrain" connectome and integrating this information with the anatomical and physiological recording of neurons in other species, we observe neuropil and lineage-specific innervation, connectivity, and activity profiles in CX input channels. We posit that the proliferative potential of neuroblasts and the lineage-based architecture of information channels enable the modification of neural networks across existing, novel, and deprecated modalities in a species-specific manner, thus forming the substrate for the evolution and diversification of insect navigational circuits.
Keywords: Central complex; Hemibrain; Insect brains; Large-field neurons; Lineages.
© 2023. The Author(s).
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as potential conflicts of interest.
Figures


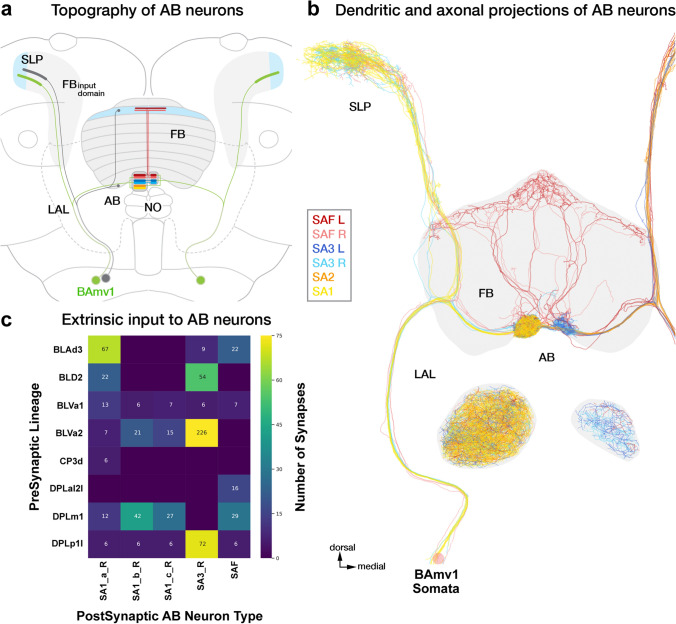
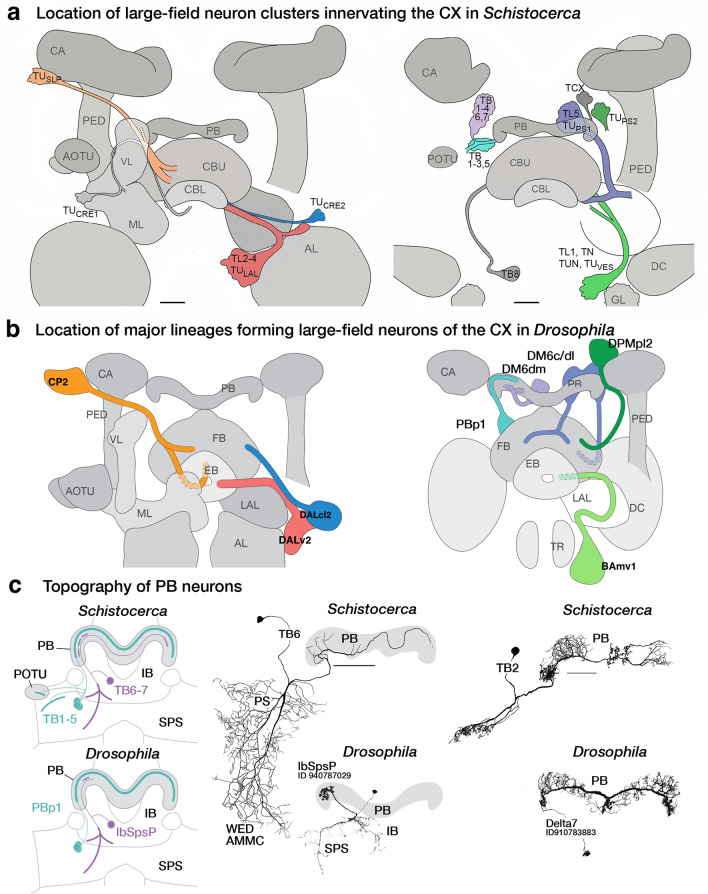
Similar articles
-
A projectome of the bumblebee central complex.Elife. 2021 Sep 15;10:e68911. doi: 10.7554/eLife.68911. Elife. 2021. PMID: 34523418 Free PMC article.
-
Drosophila type II neuroblast lineages keep Prospero levels low to generate large clones that contribute to the adult brain central complex.Neural Dev. 2010 Oct 1;5:26. doi: 10.1186/1749-8104-5-26. Neural Dev. 2010. PMID: 20920301 Free PMC article.
-
Arborization pattern of engrailed-positive neural lineages reveal neuromere boundaries in the Drosophila brain neuropil.J Comp Neurol. 2009 Nov 1;517(1):87-104. doi: 10.1002/cne.22112. J Comp Neurol. 2009. PMID: 19711412 Free PMC article.
-
Flexible navigational computations in the Drosophila central complex.Curr Opin Neurobiol. 2022 Apr;73:102514. doi: 10.1016/j.conb.2021.12.001. Epub 2022 Feb 20. Curr Opin Neurobiol. 2022. PMID: 35196623 Review.
-
Wiring the Drosophila Brain with Individually Tailored Neural Lineages.Curr Biol. 2017 Jan 23;27(2):R77-R82. doi: 10.1016/j.cub.2016.12.026. Curr Biol. 2017. PMID: 28118595 Review.
Cited by
-
From bristle to brain: embryonic development of topographic projections from basiconic sensilla in the antennal nervous system of the locust Schistocerca gregaria.Dev Genes Evol. 2024 Jun;234(1):33-44. doi: 10.1007/s00427-024-00716-2. Epub 2024 May 1. Dev Genes Evol. 2024. PMID: 38691194 Free PMC article.
-
Neuroarchitecture of the Central Complex in the Madeira Cockroach Rhyparobia maderae: Tangential Neurons.J Comp Neurol. 2024 Dec;532(12):e70009. doi: 10.1002/cne.70009. J Comp Neurol. 2024. PMID: 39658819 Free PMC article.
-
Neural circuits for goal-directed navigation across species.Trends Neurosci. 2024 Nov;47(11):904-917. doi: 10.1016/j.tins.2024.09.005. Epub 2024 Oct 10. Trends Neurosci. 2024. PMID: 39393938 Review.
-
Unraveling the neural basis of spatial orientation in arthropods.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2023 Jul;209(4):459-464. doi: 10.1007/s00359-023-01635-9. Epub 2023 May 17. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2023. PMID: 37198448 Free PMC article.
-
Neurotransmitter classification from electron microscopy images at synaptic sites in Drosophila melanogaster.Cell. 2024 May 9;187(10):2574-2594.e23. doi: 10.1016/j.cell.2024.03.016. Cell. 2024. PMID: 38729112 Free PMC article.
References
-
- Aleman A, Omoto JJ, Singh P, Nguyen B-C, Kandimalla P, Hartenstein V, Donlea JM. Opposing subclasses of Drosophila ellipsoid body neurons promote and suppress sleep. bioRxiv. 2021 doi: 10.1101/2021.10.19.464469. - DOI
-
- Beetz MJ, el Jundi B, Heinze S, Homberg U. Topographic organization and possible function of the posterior optic tubercles in the brain of the desert locust Schistocerca gregaria: neural organization of the posterior optic tubercle. J Comp Neurol. 2015;523:1589–1607. doi: 10.1002/cne.23736. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
