

SAMHD1 restricts the deoxyguanosine triphosphate pool contributing to telomere stability in telomerase-positive cells
- PMID: 36934410
- PMCID: PMC11977530
- DOI: 10.1096/fj.202300122R
SAMHD1 restricts the deoxyguanosine triphosphate pool contributing to telomere stability in telomerase-positive cells
Abstract
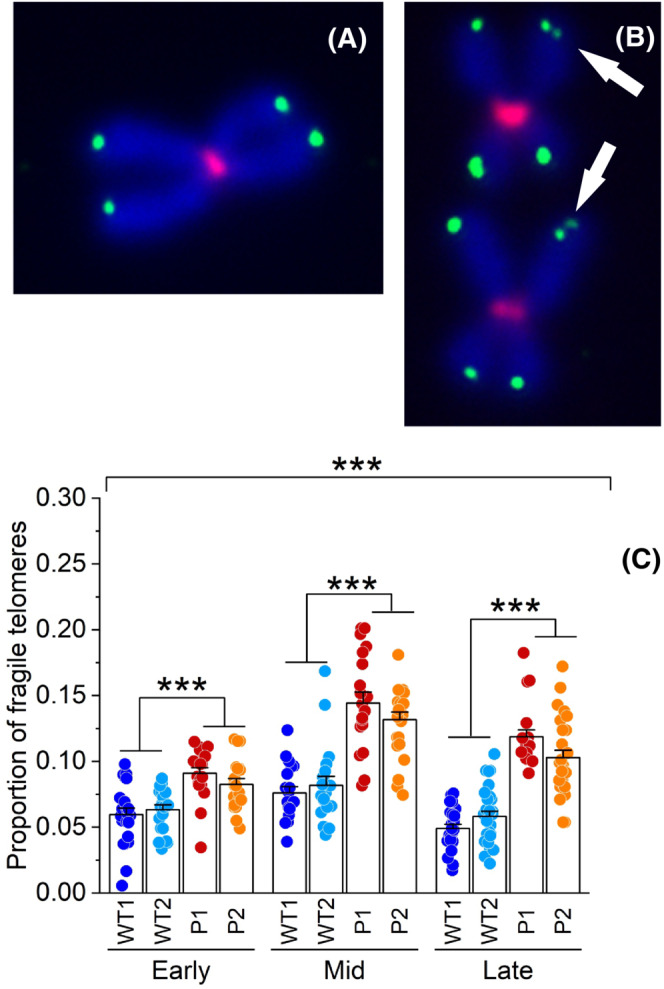
SAMHD1 (Sterile alpha motif and histidine/aspartic acid domain-containing protein 1) is a dNTP triphosphohydrolase crucial in the maintenance of balanced cellular dNTP pools, which support genome integrity. In SAMHD1 deficient fibroblasts isolated from Aicardi-Goutières Syndrome (AGS) patients, all four DNA precursors are increased and markedly imbalanced with the largest effect on dGTP, a key player in the modulation of telomerase processivity. Here, we present data showing that SAMHD1, by restricting the dGTP pool, contributes to telomere maintenance in hTERT-immortalized human fibroblasts from AGS patients as well as in telomerase positive cancer cell lines. Only in cells expressing telomerase, the lack of SAMHD1 causes excessive lengthening of telomeres and telomere fragility, whereas primary fibroblasts lacking both SAMHD1 and telomerase enter normally into senescence. Telomere lengthening observed in SAMHD1 deficient but telomerase proficient cells is a gradual process, in accordance with the intrinsic property of telomerase of adding only a few tens of nucleotides for each cycle. Therefore, only a prolonged exposure to high dGTP content causes telomere over-elongation. hTERT-immortalized AGS fibroblasts display also high fragility of chromosome ends, a marker of telomere replication stress. These results not only demonstrate the functional importance of dGTP cellular level but also reveal the critical role played by SAMHD1 in restraining telomerase processivity and safeguarding telomere stability.
Keywords: Aicardi-Goutières syndrome (AGS); deoxynucleotide metabolism; deoxynucleotide pool imbalance; sterile alpha motif and HD domain containing protein 1 (SAMHD1); telomere maintenance.
© 2023 The Authors. The FASEB Journal published by Wiley Periodicals LLC on behalf of Federation of American Societies for Experimental Biology.
Figures





References
-
- Goldstone DC, Ennis‐Adeniran V, Hedden JJ, et al. HIV‐1 restriction factor SAMHD1 is a deoxynucleoside triphosphate triphosphohydrolase. Nature. 2011;480:379‐382. - PubMed
-
- Coquel F, Silva M‐J, Técher H, et al. SAMHD1 acts at stalled replication forks to prevent interferon induction. Nature. 2018;557:57‐61. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
