

Proteasome inhibitor bortezomib stabilizes and activates p53 in hematopoietic stem/progenitors and double-negative T cells in vivo
- PMID: 36940336
- PMCID: PMC10068759
- DOI: 10.1073/pnas.2219978120
Proteasome inhibitor bortezomib stabilizes and activates p53 in hematopoietic stem/progenitors and double-negative T cells in vivo
Abstract
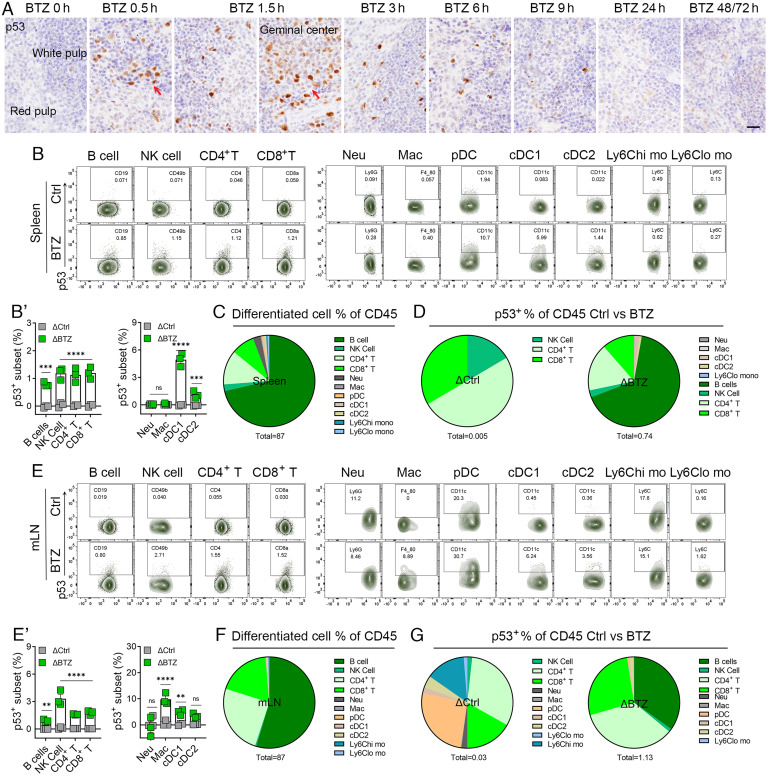
We have previously shown that proteasome inhibitor bortezomib stabilizes p53 in stem and progenitor cells within gastrointestinal tissues. Here, we characterize the effect of bortezomib treatment on primary and secondary lymphoid tissues in mice. We find that bortezomib stabilizes p53 in significant fractions of hematopoietic stem and progenitor cells in the bone marrow, including common lymphoid and myeloid progenitors, granulocyte-monocyte progenitors, and dendritic cell progenitors. The stabilization of p53 is also observed in multipotent progenitors and hematopoietic stem cells, albeit at lower frequencies. In the thymus, bortezomib stabilizes p53 in CD4-CD8- T cells. Although there is less p53 stabilization in secondary lymphoid organs, cells in the germinal center of the spleen and Peyer's patch accumulate p53 in response to bortezomib. Bortezomib induces the upregulation of p53 target genes and p53 dependent/independent apoptosis in the bone marrow and thymus, suggesting that cells in these organs are robustly affected by proteasome inhibition. Comparative analysis of cell percentages in the bone marrow indicates expanded stem and multipotent progenitor pools in p53R172H mutant mice compared with p53 wild-type mice, suggesting a critical role for p53 in regulating the development and maturation of hematopoietic cells in the bone marrow. We propose that progenitors along the hematopoietic differentiation pathway express relatively high levels of p53 protein, which under steady-state conditions is constantly degraded by Mdm2 E3 ligase; however, these cells rapidly respond to stress to regulate stem cell renewal and consequently maintain the genomic integrity of hematopoietic stem/progenitor cell populations.
Keywords: T cells; bortezomib; cancer; hematopoietic; p53.
Conflict of interest statement
The authors declare no competing interest.
Figures
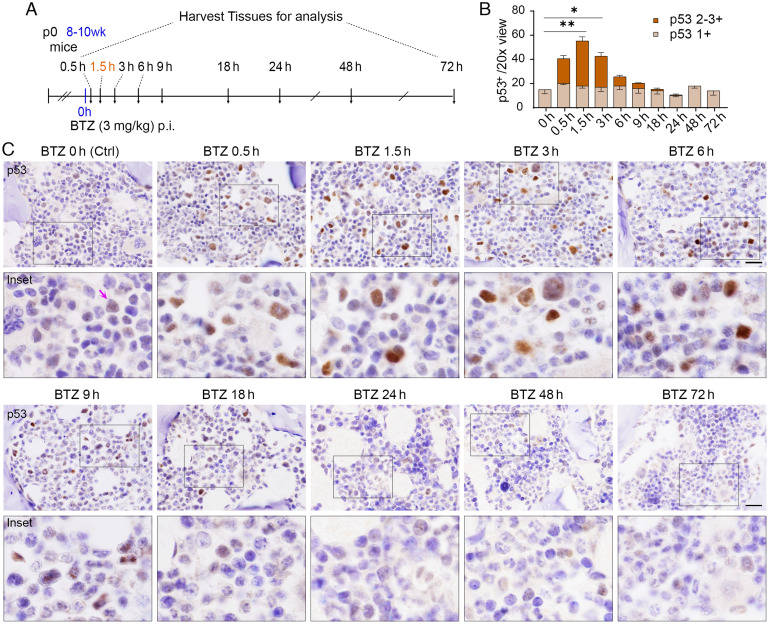
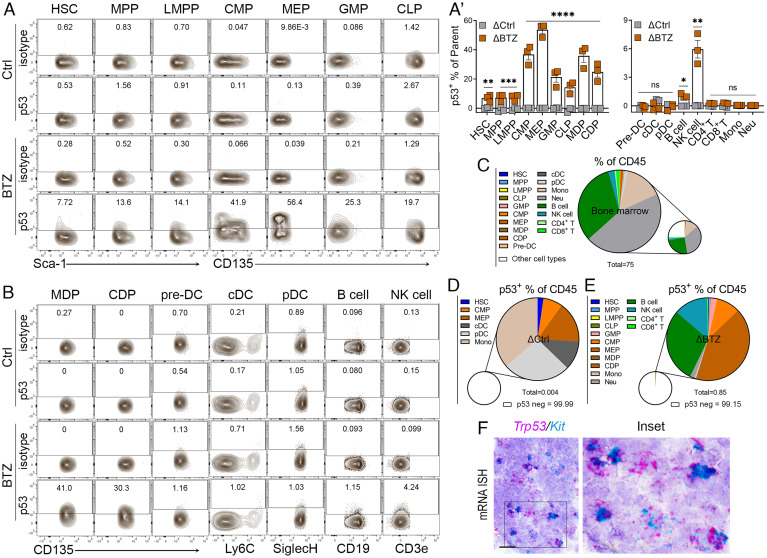
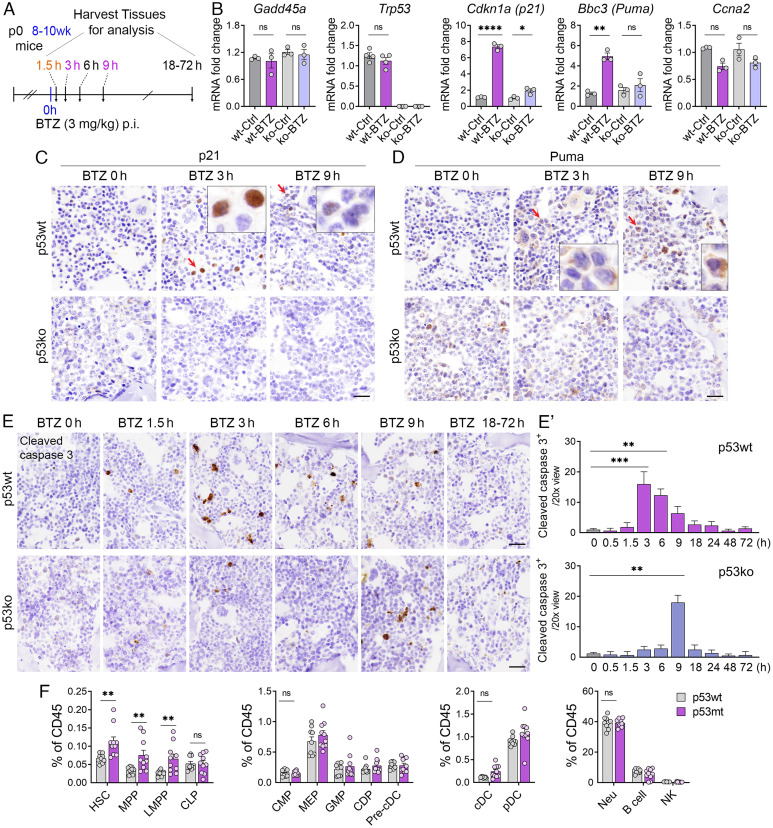
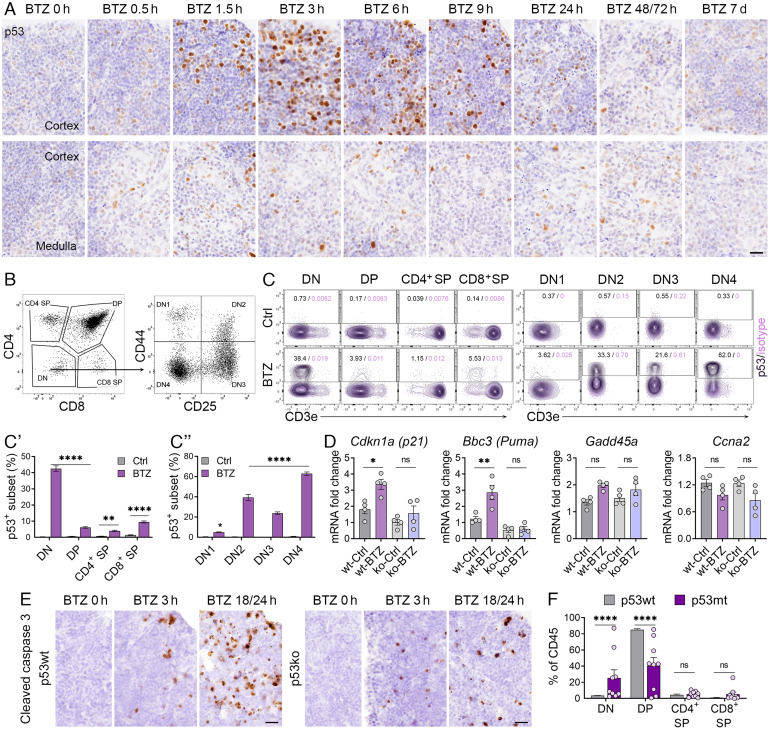
References
-
- Jacobsen S. E. W., Nerlov C., Haematopoiesis in the era of advanced single-cell technologies. Nat. Cell Biol. 21, 2–8 (2019). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
