

Shaping the brain vasculature in development and disease in the single-cell era
- PMID: 36941369
- PMCID: PMC10026800
- DOI: 10.1038/s41583-023-00684-y
Shaping the brain vasculature in development and disease in the single-cell era
Abstract
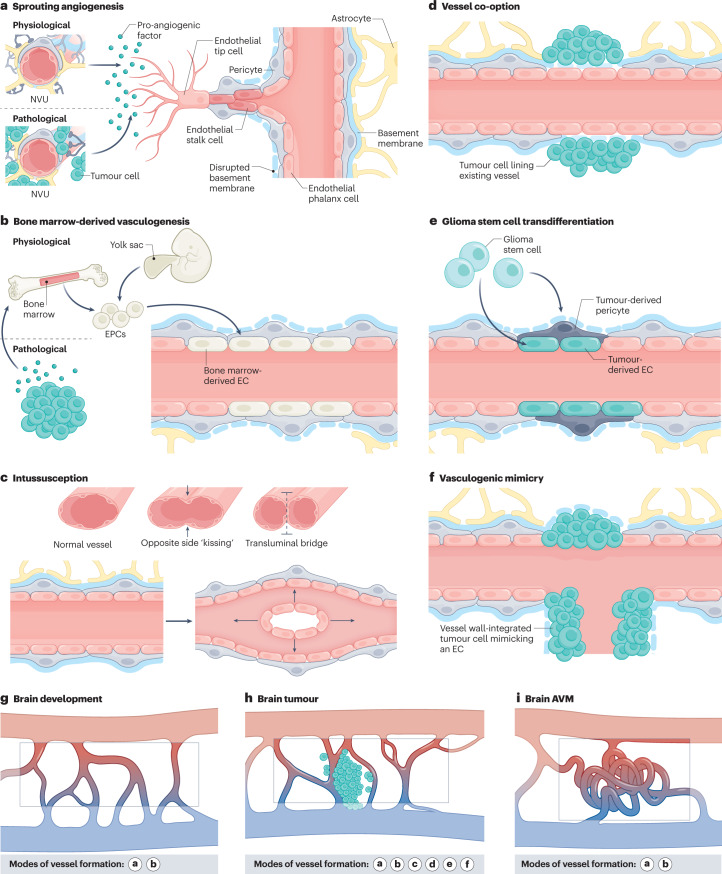
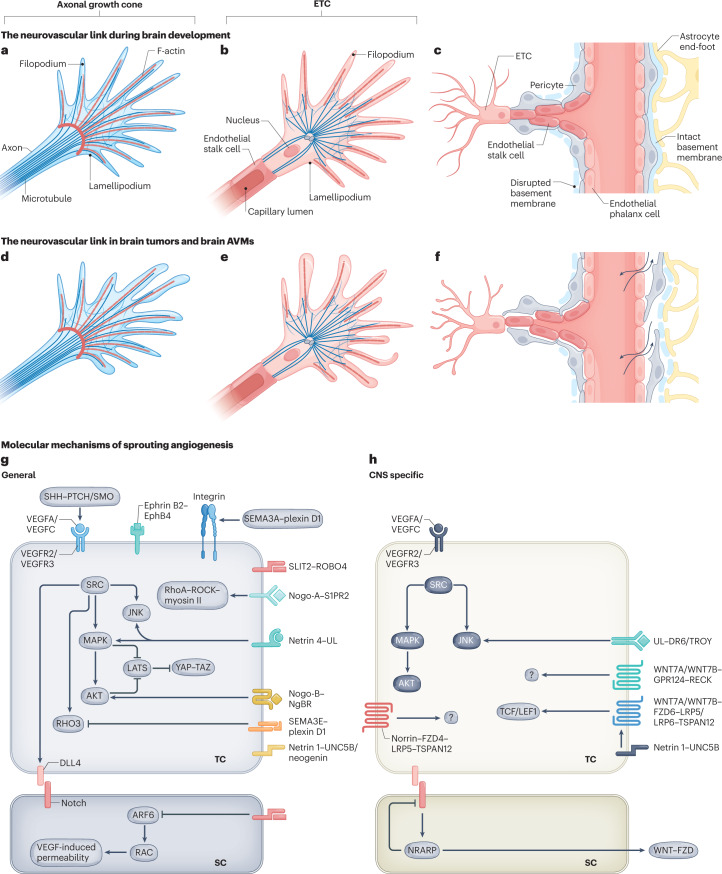
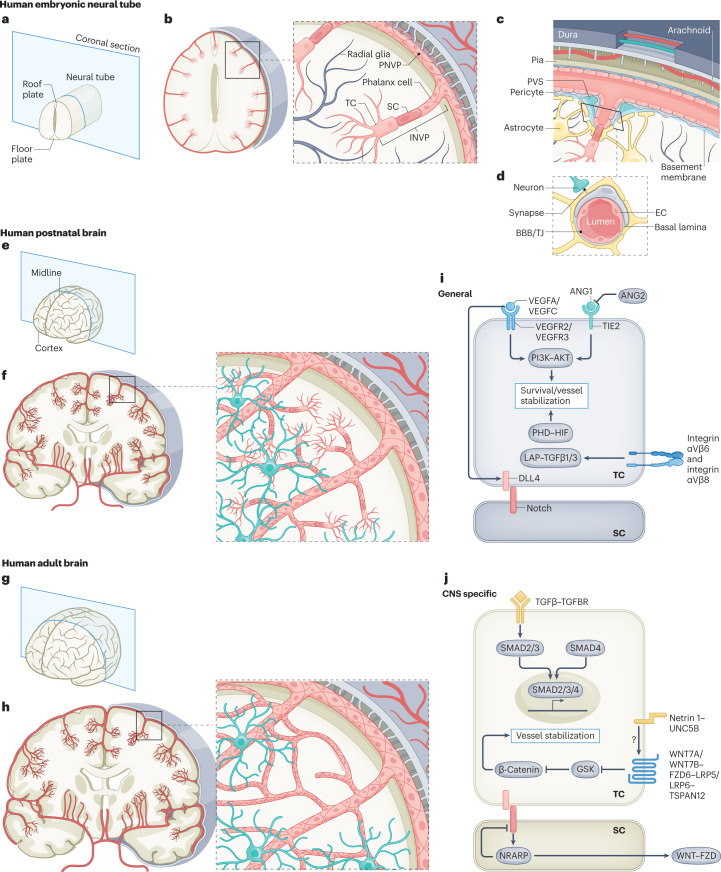
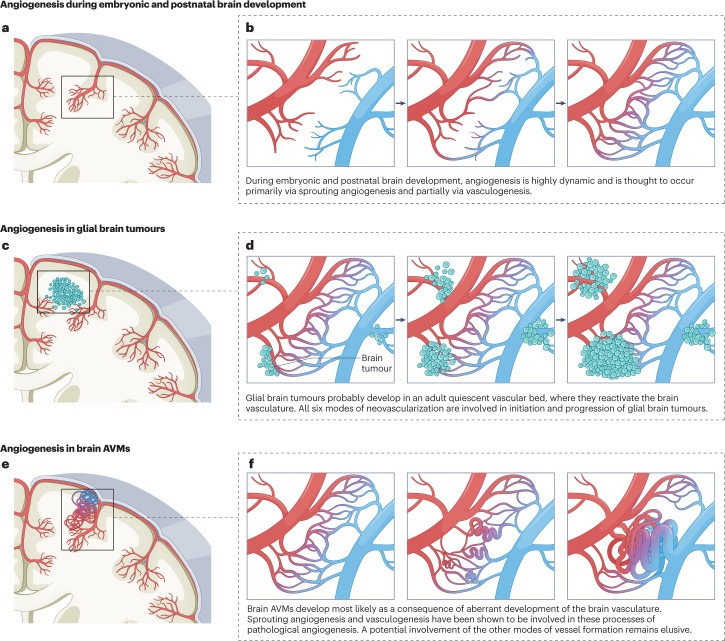
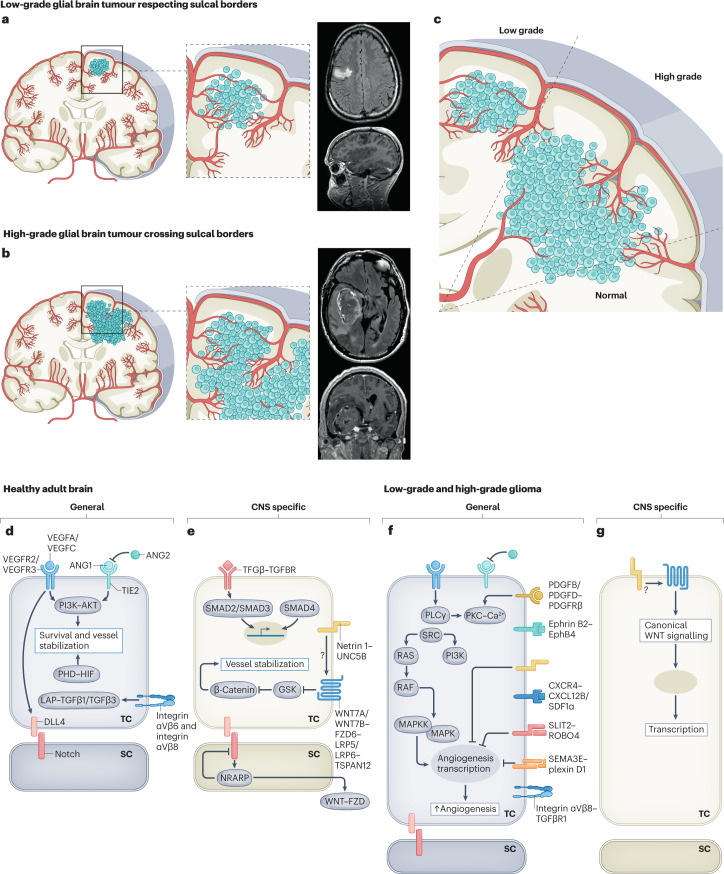
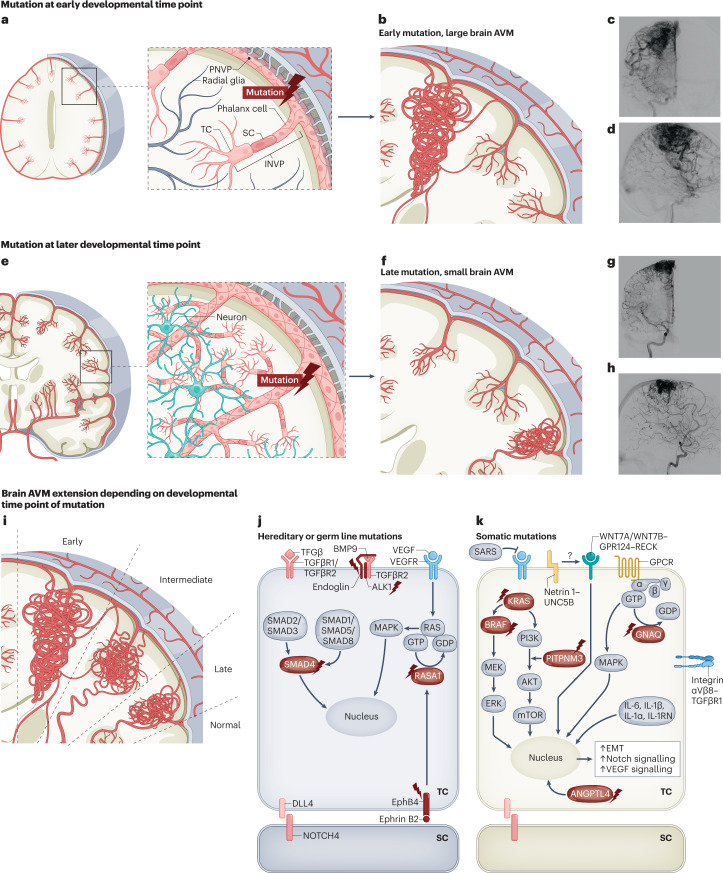
The CNS critically relies on the formation and proper function of its vasculature during development, adult homeostasis and disease. Angiogenesis - the formation of new blood vessels - is highly active during brain development, enters almost complete quiescence in the healthy adult brain and is reactivated in vascular-dependent brain pathologies such as brain vascular malformations and brain tumours. Despite major advances in the understanding of the cellular and molecular mechanisms driving angiogenesis in peripheral tissues, developmental signalling pathways orchestrating angiogenic processes in the healthy and the diseased CNS remain incompletely understood. Molecular signalling pathways of the 'neurovascular link' defining common mechanisms of nerve and vessel wiring have emerged as crucial regulators of peripheral vascular growth, but their relevance for angiogenesis in brain development and disease remains largely unexplored. Here we review the current knowledge of general and CNS-specific mechanisms of angiogenesis during brain development and in brain vascular malformations and brain tumours, including how key molecular signalling pathways are reactivated in vascular-dependent diseases. We also discuss how these topics can be studied in the single-cell multi-omics era.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Mink JW, Blumenschine RJ, Adams DB. Ratio of central nervous system to body metabolism in vertebrates: its constancy and functional basis. Am. J. Physiol. 1981;241:R203–R212. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
