

Surviving and Adapting to Stress: Translational Control and the Integrated Stress Response
- PMID: 36943285
- PMCID: PMC10443206
- DOI: 10.1089/ars.2022.0123
Surviving and Adapting to Stress: Translational Control and the Integrated Stress Response
Abstract
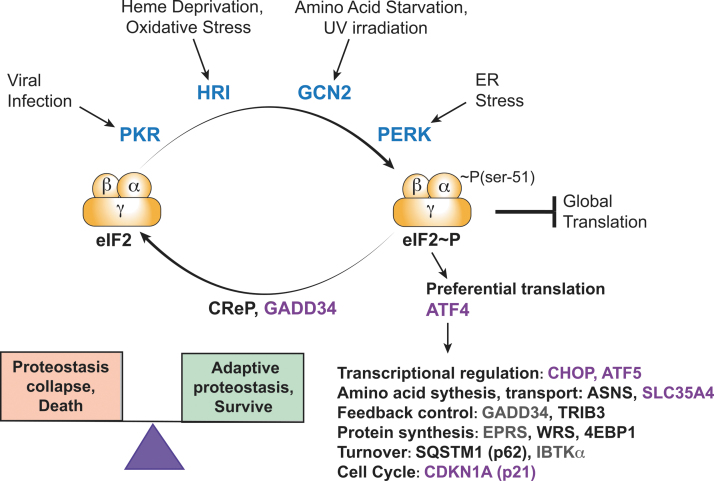
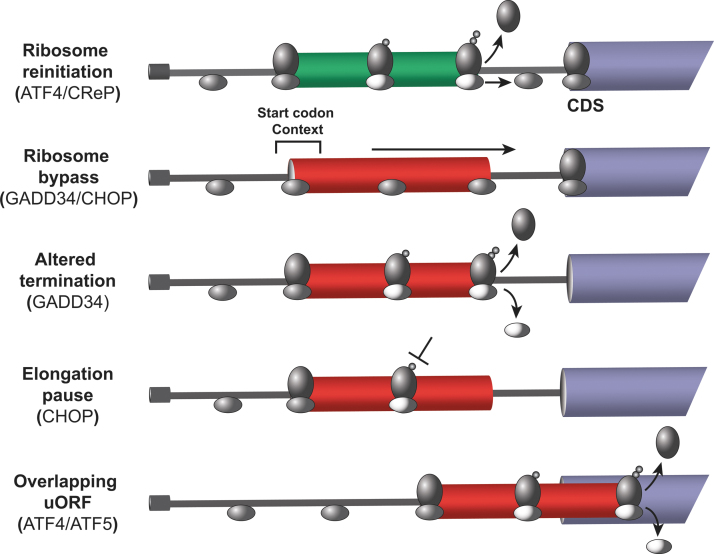
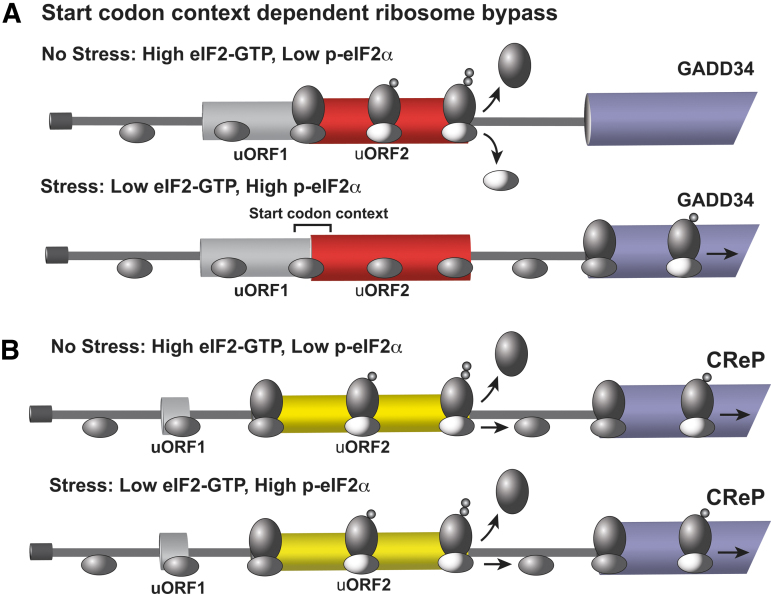
Significance: Organisms adapt to changing environments by engaging cellular stress response pathways that serve to restore proteostasis and enhance survival. A primary adaptive mechanism is the integrated stress response (ISR), which features phosphorylation of the α subunit of eukaryotic translation initiation factor 2 (eIF2). Four eIF2α kinases respond to different stresses, enabling cells to rapidly control translation to optimize management of resources and reprogram gene expression for stress adaptation. Phosphorylation of eIF2 blocks its guanine nucleotide exchange factor, eIF2B, thus lowering the levels of eIF2 bound to GTP that is required to deliver initiator transfer RNA (tRNA) to ribosomes. While bulk messenger RNA (mRNA) translation can be sharply lowered by heightened phosphorylation of eIF2α, there are other gene transcripts whose translation is unchanged or preferentially translated. Among the preferentially translated genes is ATF4, which directs transcription of adaptive genes in the ISR. Recent Advances and Critical Issues: This review focuses on how eIF2α kinases function as first responders of stress, the mechanisms by which eIF2α phosphorylation and other stress signals regulate the exchange activity of eIF2B, and the processes by which the ISR triggers differential mRNA translation. To illustrate the synergy between stress pathways, we describe the mechanisms and functional significance of communication between the ISR and another key regulator of translation, mammalian/mechanistic target of rapamycin complex 1 (mTORC1), during acute and chronic amino acid insufficiency. Finally, we discuss the pathological conditions that stem from aberrant regulation of the ISR, as well as therapeutic strategies targeting the ISR to alleviate disease. Future Directions: Important topics for future ISR research are strategies for modulating this stress pathway in disease conditions and drug development, molecular processes for differential translation and the coordinate regulation of GCN2 and other stress pathways during physiological and pathological conditions. Antioxid. Redox Signal. 39, 351-373.
Keywords: eIF2; eIF2 kinases; integrated stress response; translation initiation; translational control.
Conflict of interest statement
R.C.W. is a member of the advisory board in HiberCell, Inc. K.A.S. and T.G.A. are consultants for HiberCell, Inc., and K.A.S. receives research support from HiberCell, Inc.
Figures





References
-
- Abastado JP, Miller PF, Hinnebusch AG. A quantitative model for translational control of the GCN4 gene of Saccharomyces cerevisiae. New Biol 1991a;3(5):511–524. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
