

Hedgehog-induced ZFYVE21 promotes chronic vascular inflammation by activating NLRP3 inflammasomes in T cells
- PMID: 36943921
- PMCID: PMC10061549
- DOI: 10.1126/scisignal.abo3406
Hedgehog-induced ZFYVE21 promotes chronic vascular inflammation by activating NLRP3 inflammasomes in T cells
Abstract
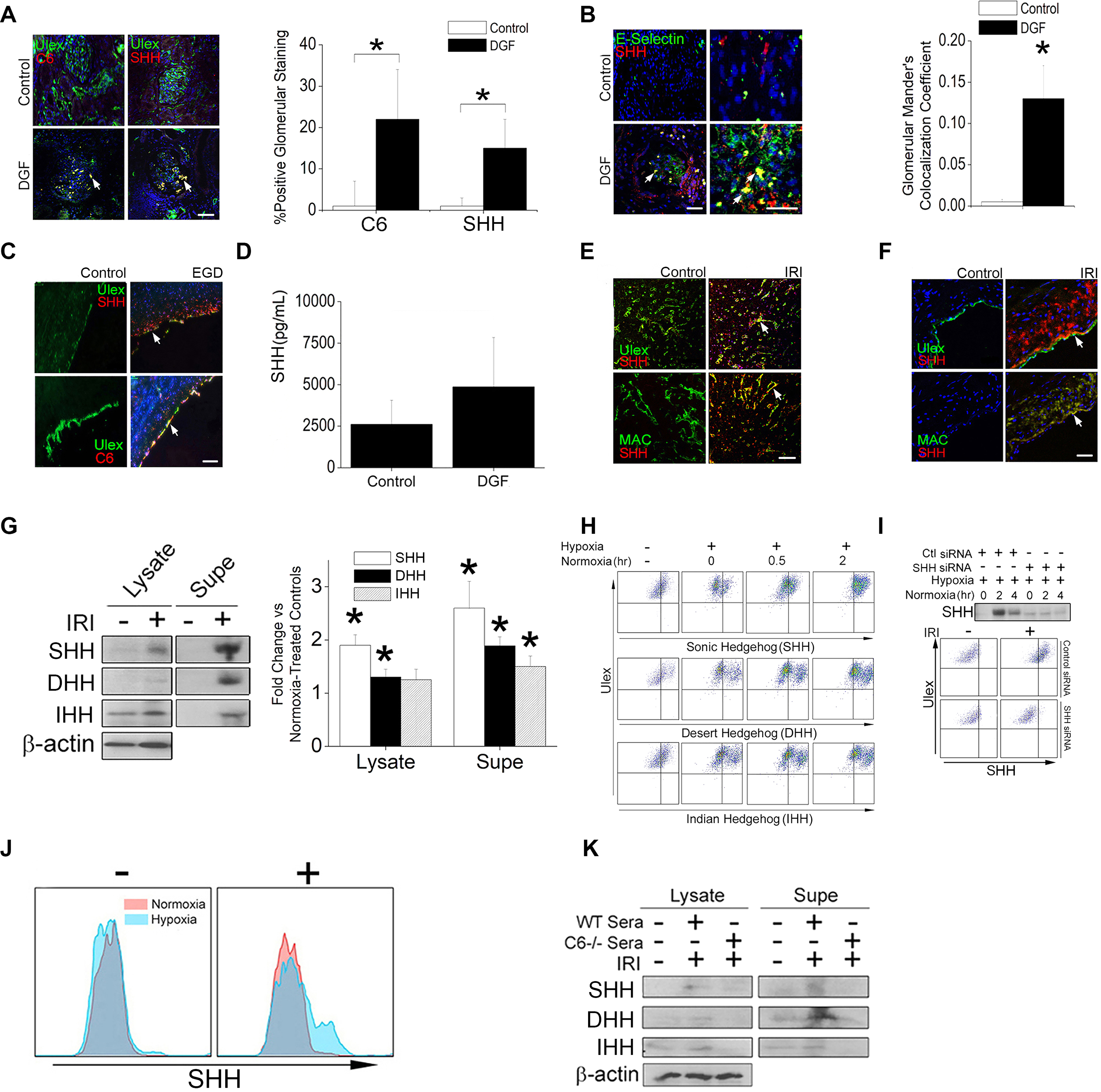
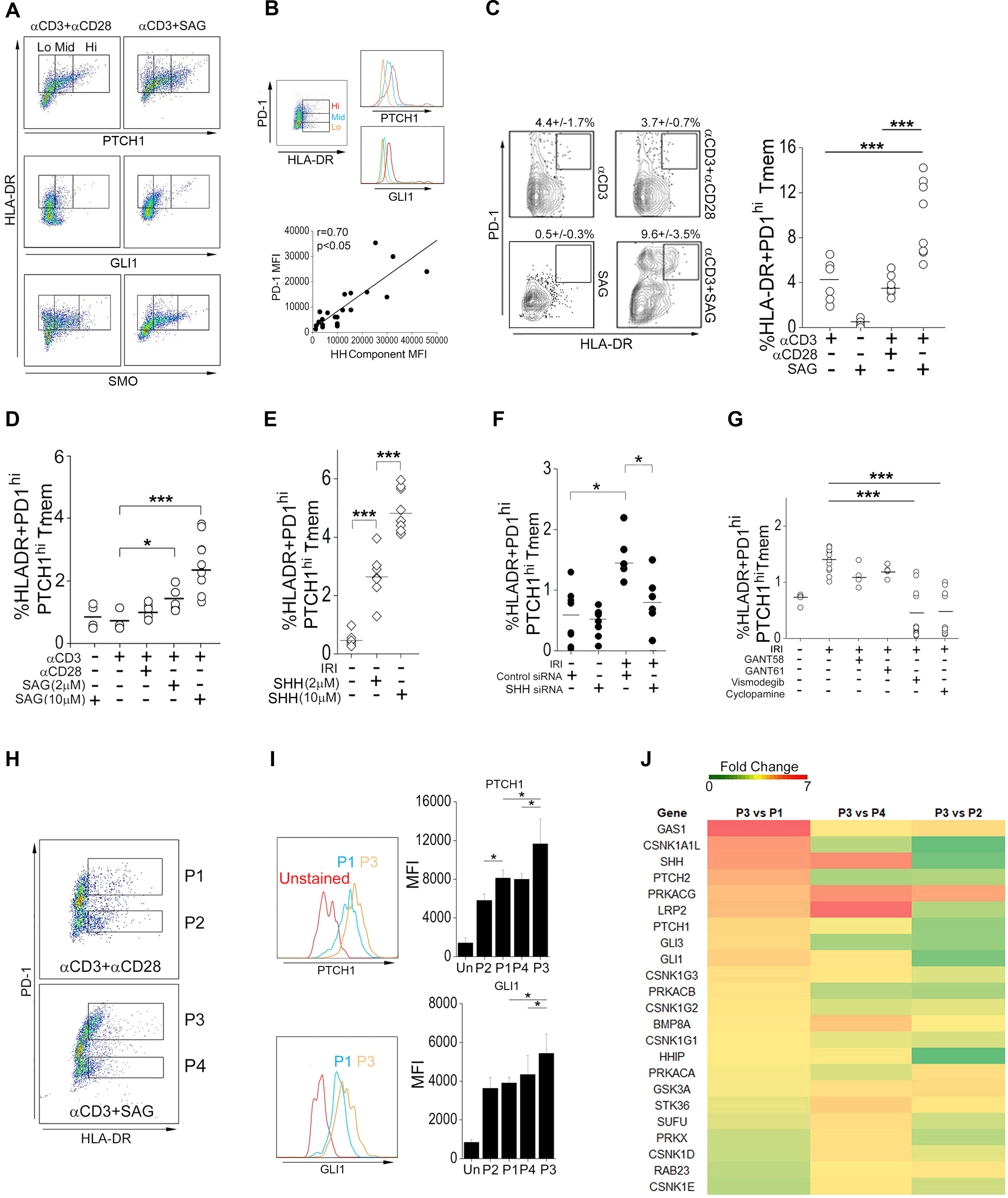
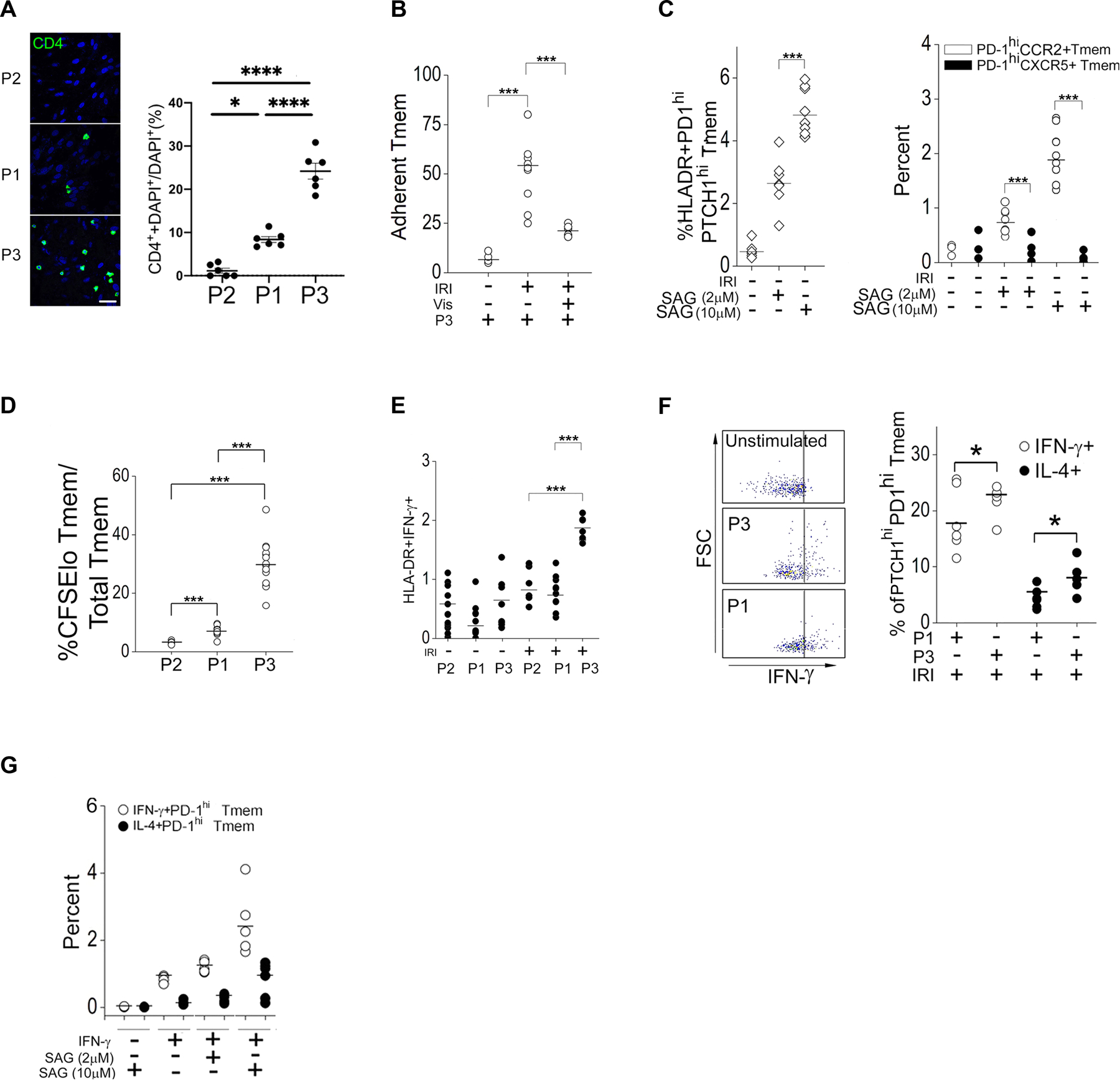
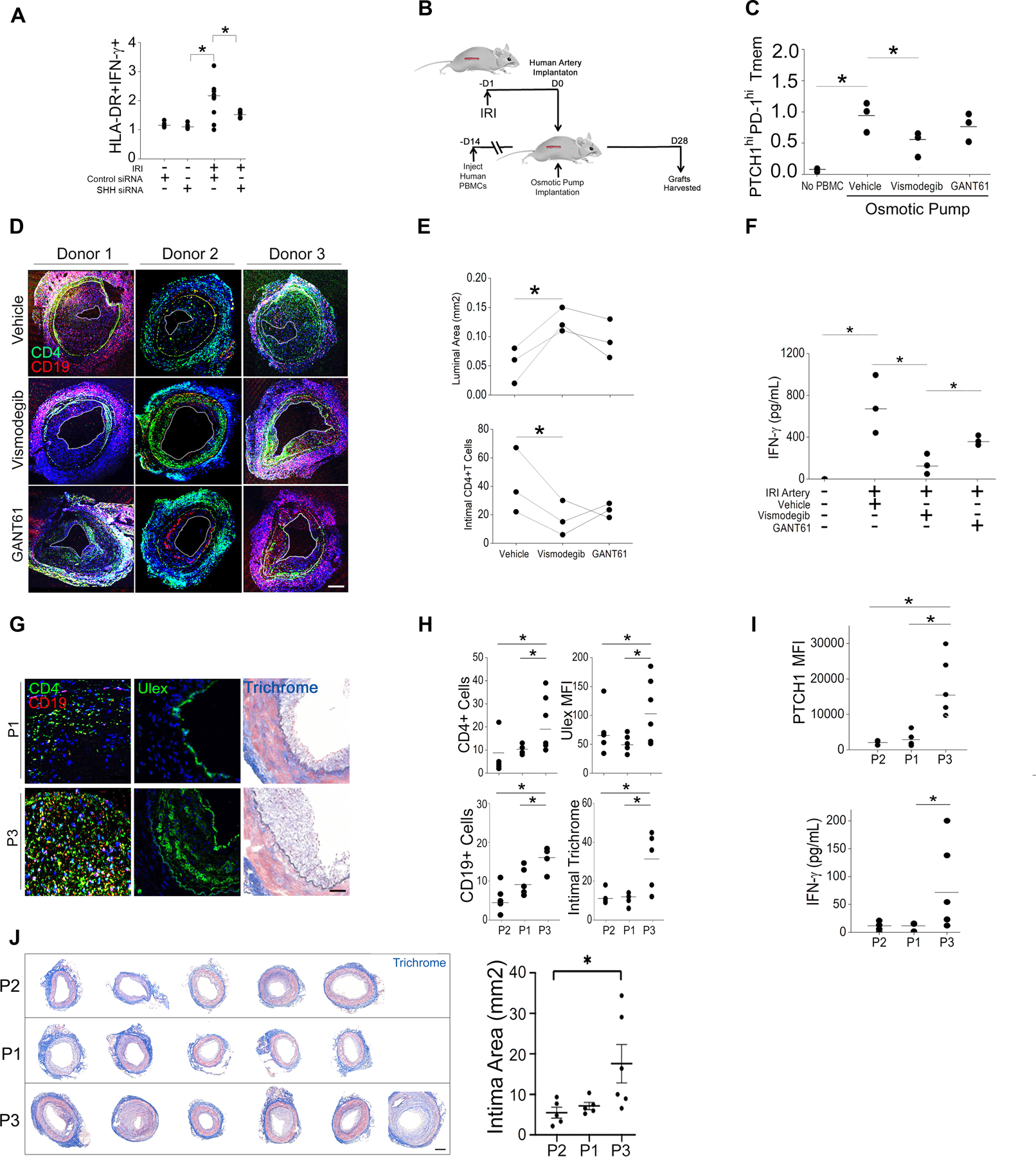
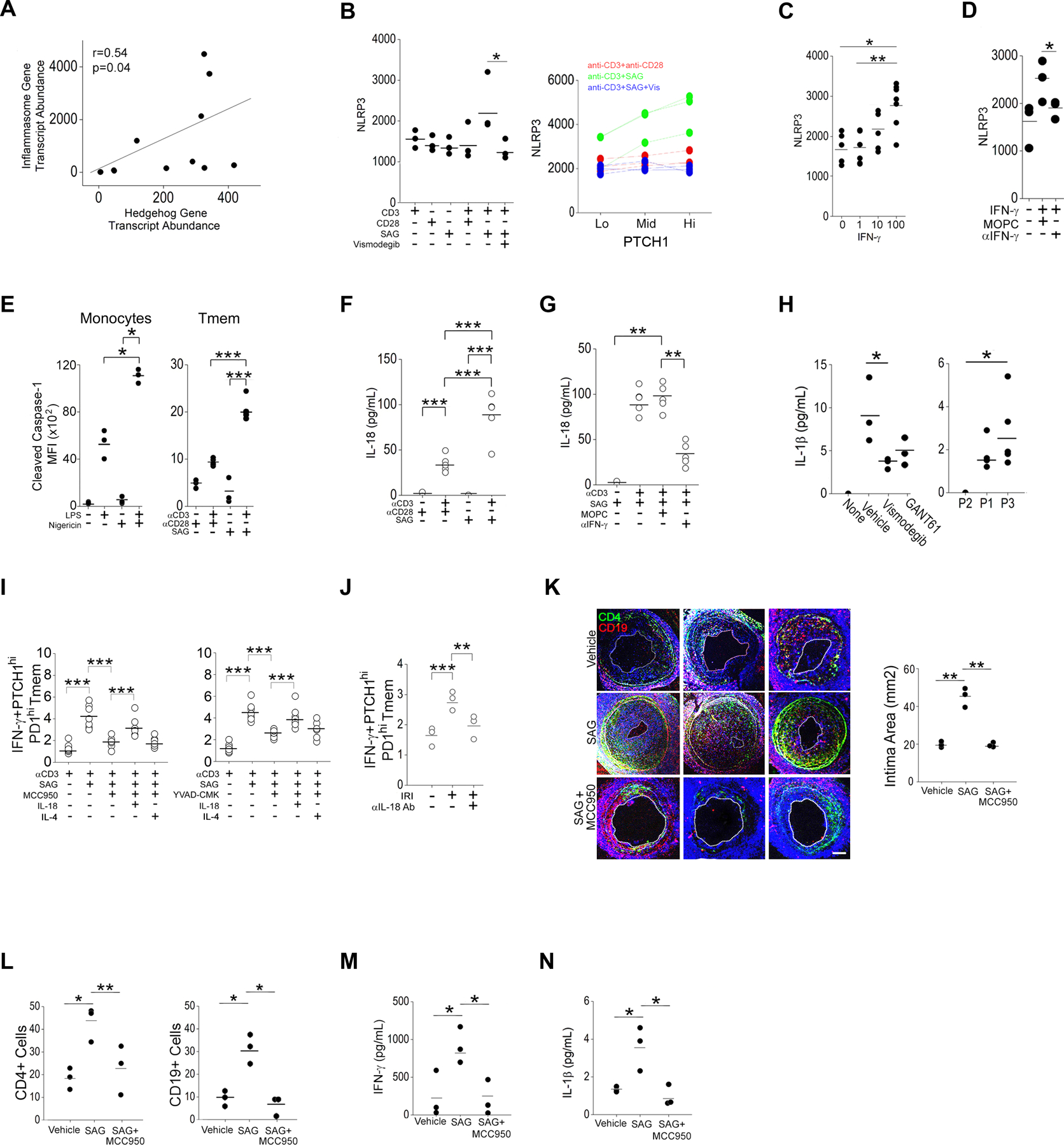
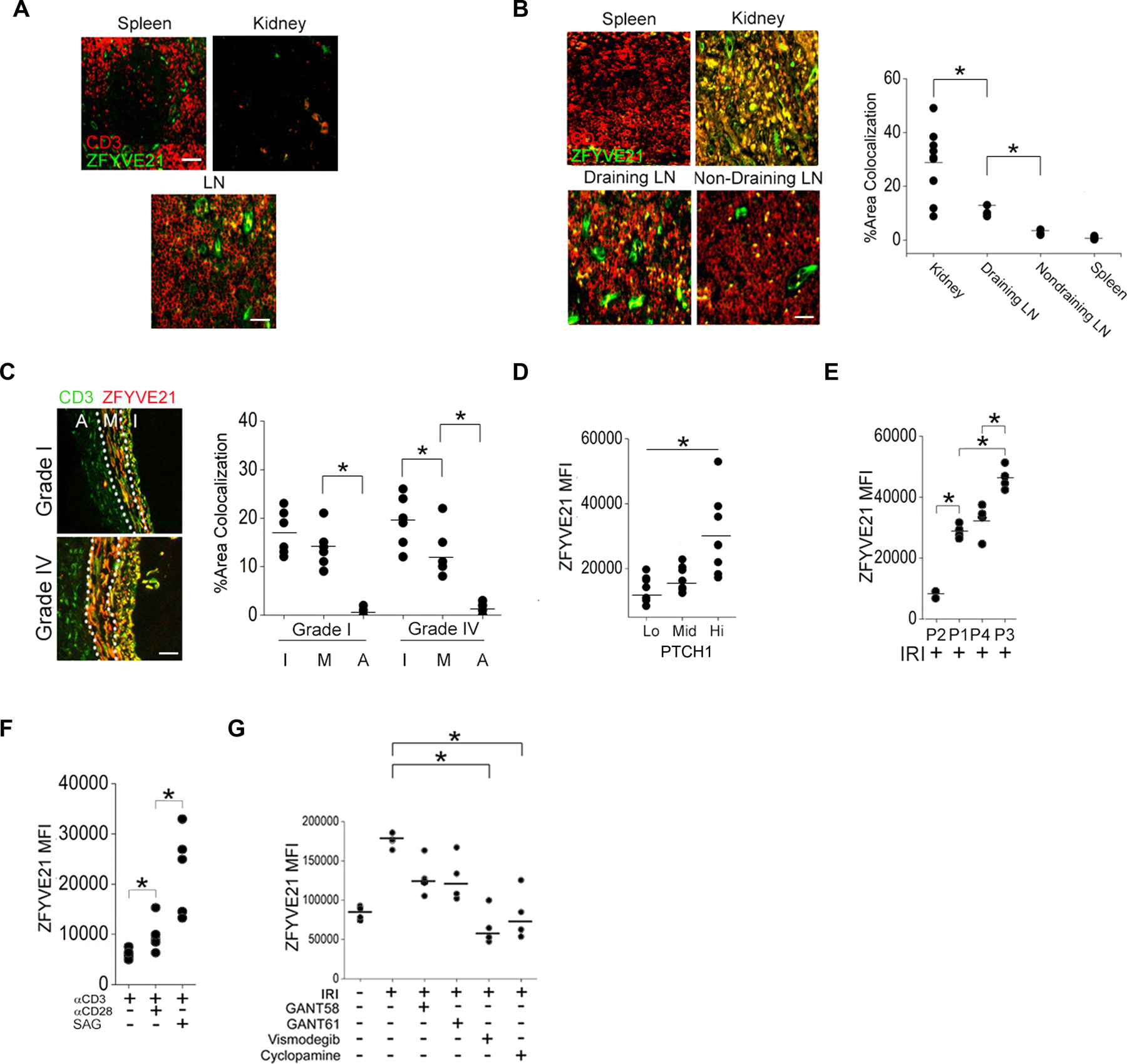
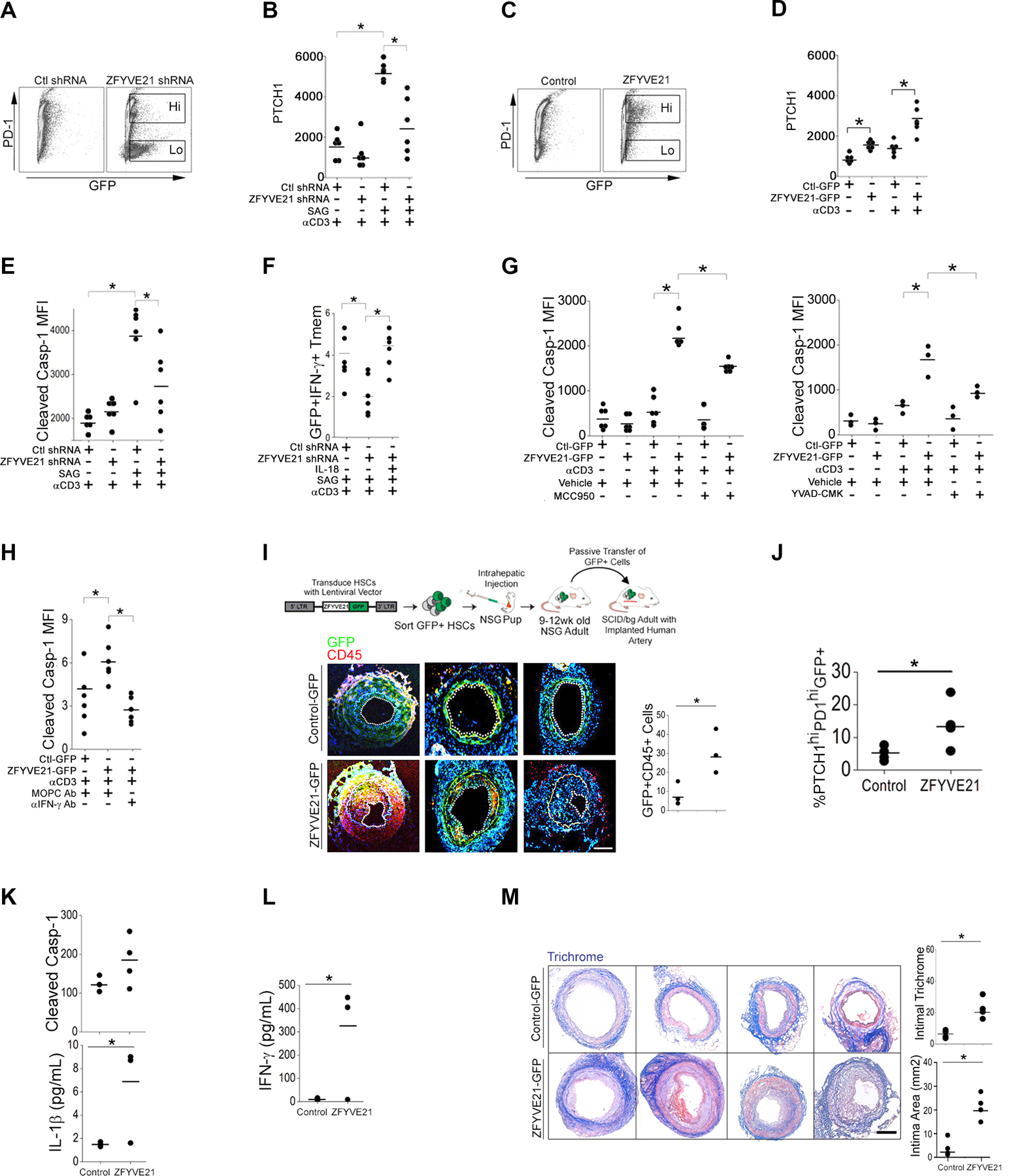
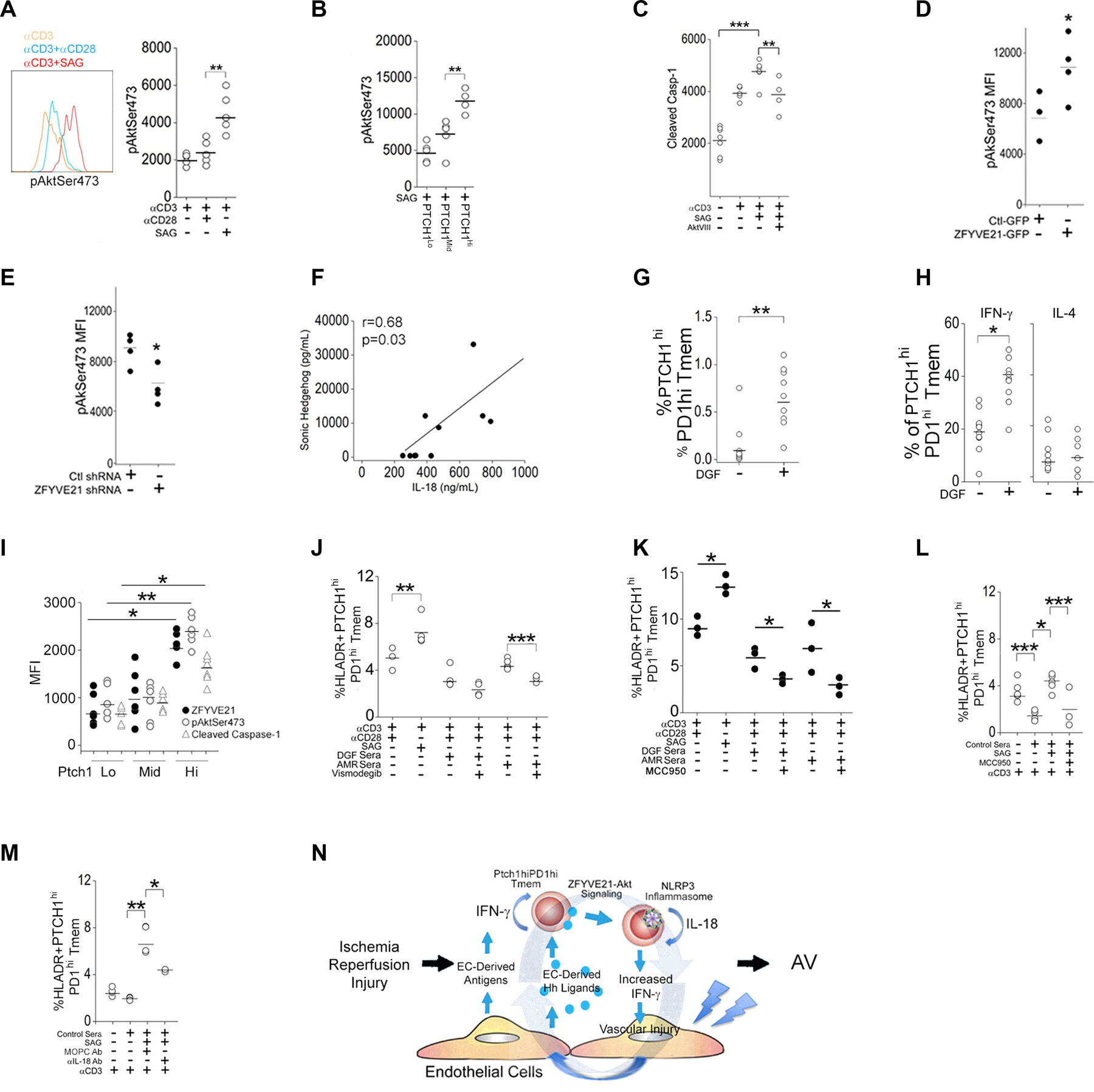
The zinc finger protein ZFYVE21 is involved in immune signaling. Using humanized mouse models, primary human cells, and patient samples, we identified a T cell-autonomous role for ZFYVE21 in promoting chronic vascular inflammation associated with allograft vasculopathy. Ischemia-reperfusion injury (IRI) stimulated endothelial cells to produce Hedgehog (Hh) ligands, which in turn induced the production of ZFYVE21 in a population of T memory cells with high amounts of the Hh receptor PTCH1 (PTCHhi cells, CD3+CD4+CD45RO+PTCH1hiPD-1hi), vigorous recruitment to injured endothelia, and increased effector responses in vivo. After priming by interferon-γ (IFN-γ), Hh-induced ZFYVE21 activated NLRP3 inflammasome activity in T cells, which potentiated IFN-γ responses. Hh-induced NLRP3 inflammasomes and T cell-specific ZFYVE21 augmented the vascular sequelae of chronic inflammation in mice engrafted with human endothelial cells or coronary arteries that had been subjected to IRI before engraftment. Moreover, the population of PTCHhi T cells producing high amounts of ZFYVE21 was expanded in patients with renal transplant-associated IRI, and sera from these patients expanded this population in control T cells in a manner that depended on Hh signaling. We conclude that Hh-induced ZFYVE21 activates NLRP3 inflammasomes in T cells, thereby promoting chronic inflammation.
Conflict of interest statement
Figures
Comment in
-
Hedgehog signalling in allograft vasculopathy: a new therapeutic target?Trends Pharmacol Sci. 2023 Sep;44(9):558-560. doi: 10.1016/j.tips.2023.05.005. Epub 2023 Jun 7. Trends Pharmacol Sci. 2023. PMID: 37296035 Free PMC article.
References
-
- Uekita T, Gotoh I, Kinoshita T, Itoh Y, Sato H, Shiomi T, Okada Y, Seiki M. Membrane-type 1 matrix metalloproteinase cytoplasmic tail-binding protein-1 is a new member of the Cupin superfamily. A possible multifunctional protein acting as an invasion suppressor down-regulated in tumors. J Biol Chem. 2004. 279:12734–43. - PubMed
-
- Nagano M, Hoshino D, Koshiba S, Shuo T, Koshikawa N, Tomizawa T, Hayashi F, Tochio N, Harada T, Akizawa T, Watanabe S, Handa N, Shirouzu M, Kigawa T, Yokoyama S, Seiki M. ZF21 protein, a regulator of the disassembly of focal adhesions and cancer metastasis, contains a novel noncanonical pleckstrin homology domain. J Biol Chem. 2011. 286:31598–609. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
