

Human T cell generation is restored in CD3δ severe combined immunodeficiency through adenine base editing
- PMID: 36944331
- PMCID: PMC10876291
- DOI: 10.1016/j.cell.2023.02.027
Human T cell generation is restored in CD3δ severe combined immunodeficiency through adenine base editing
Abstract
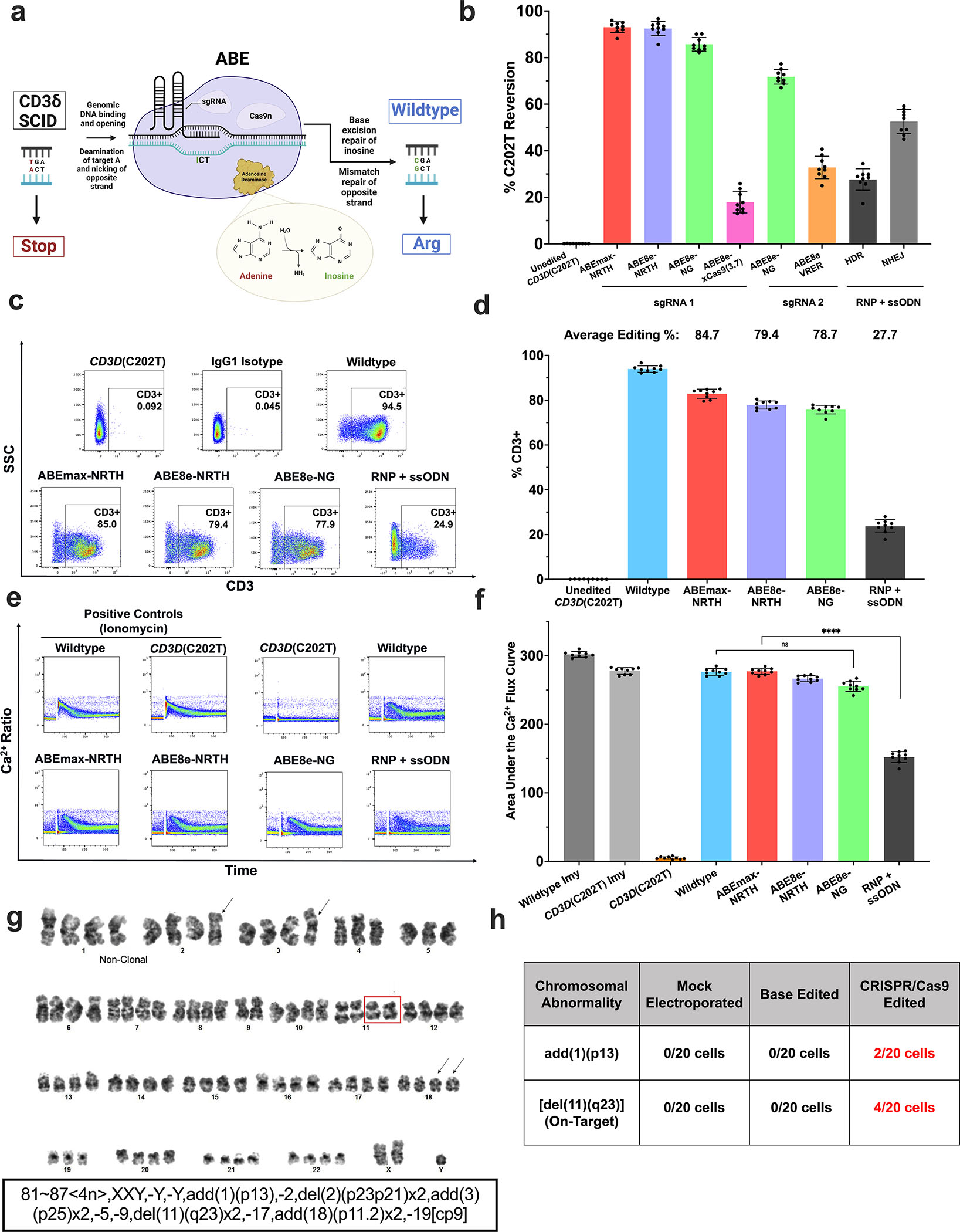
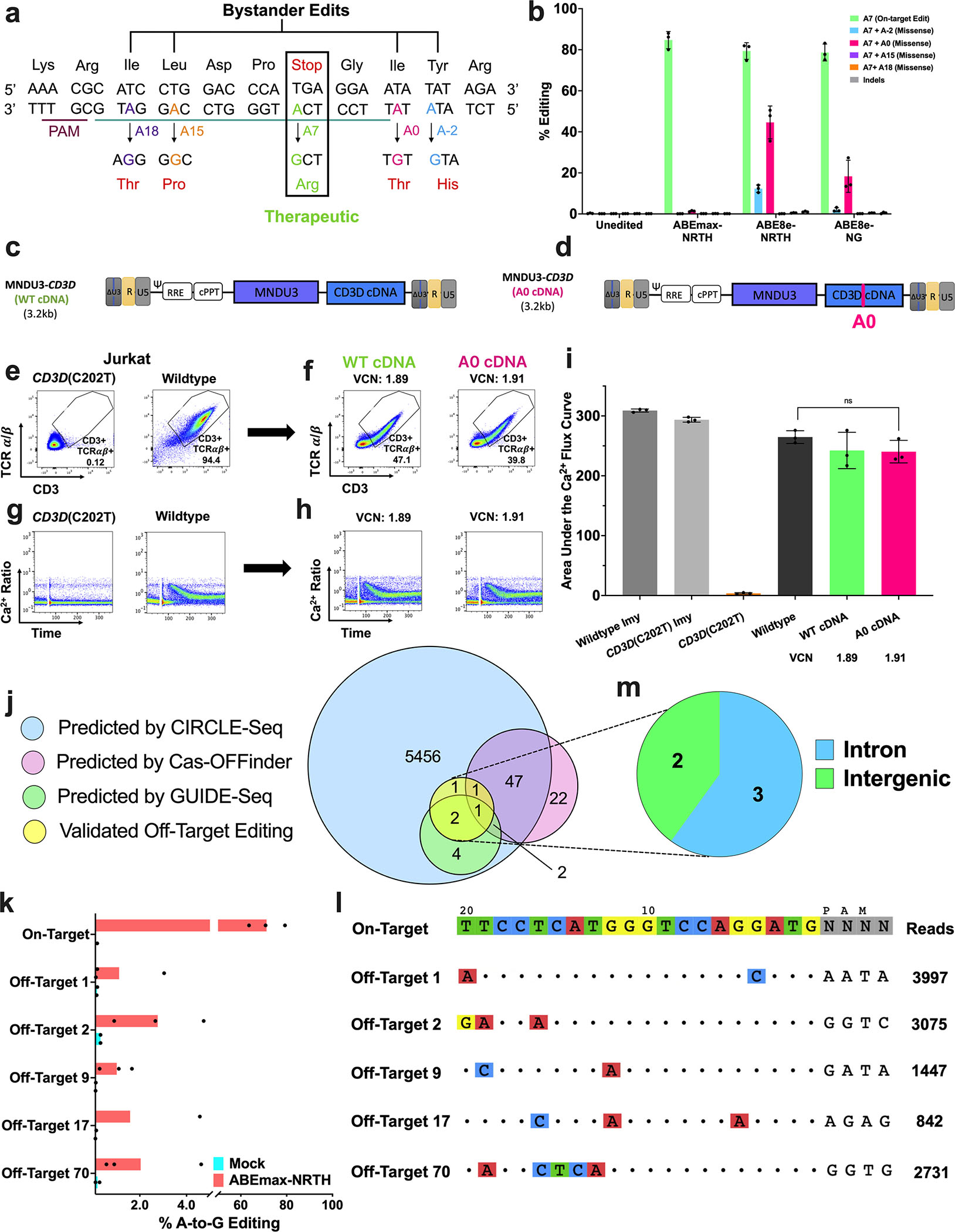
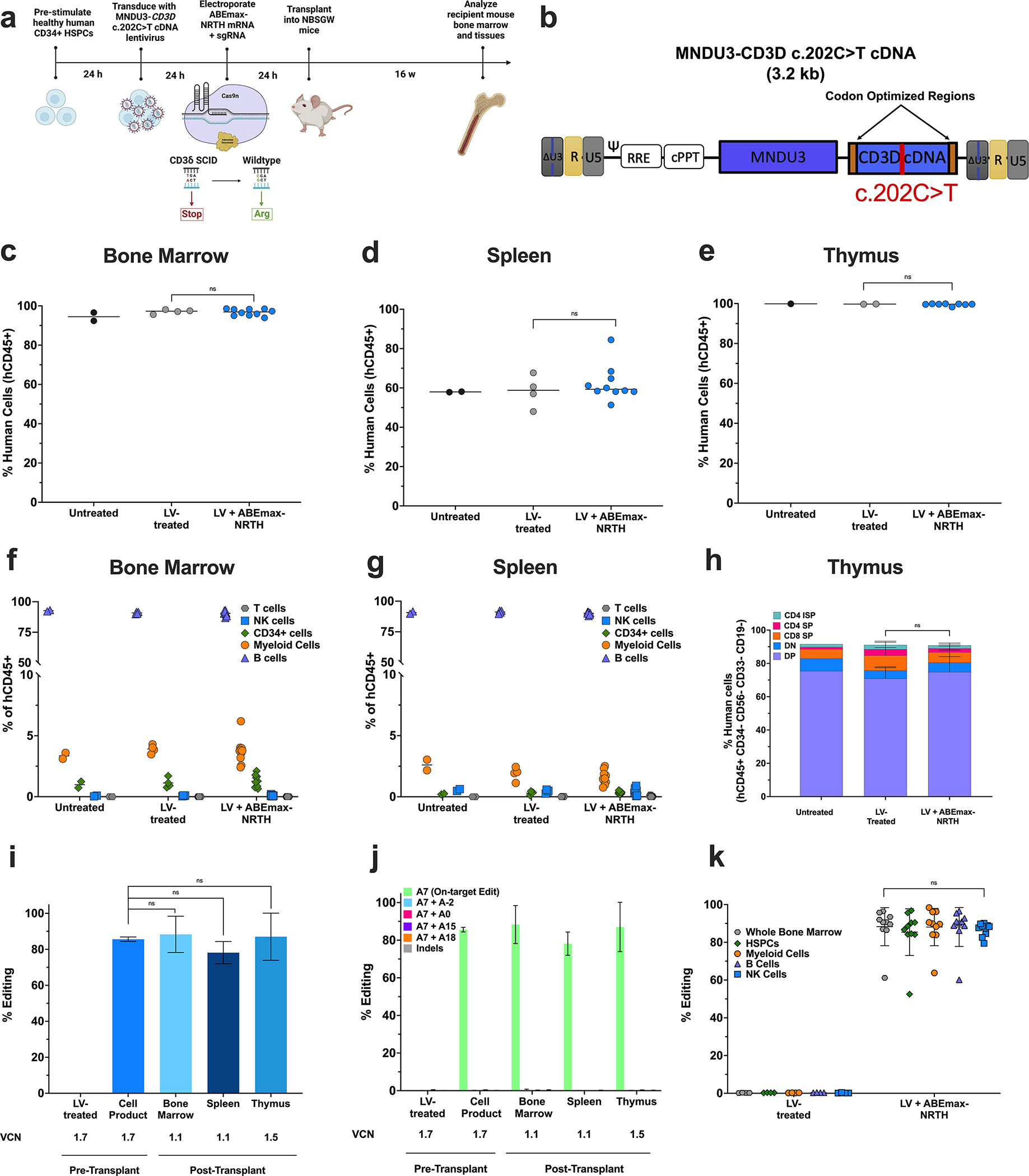
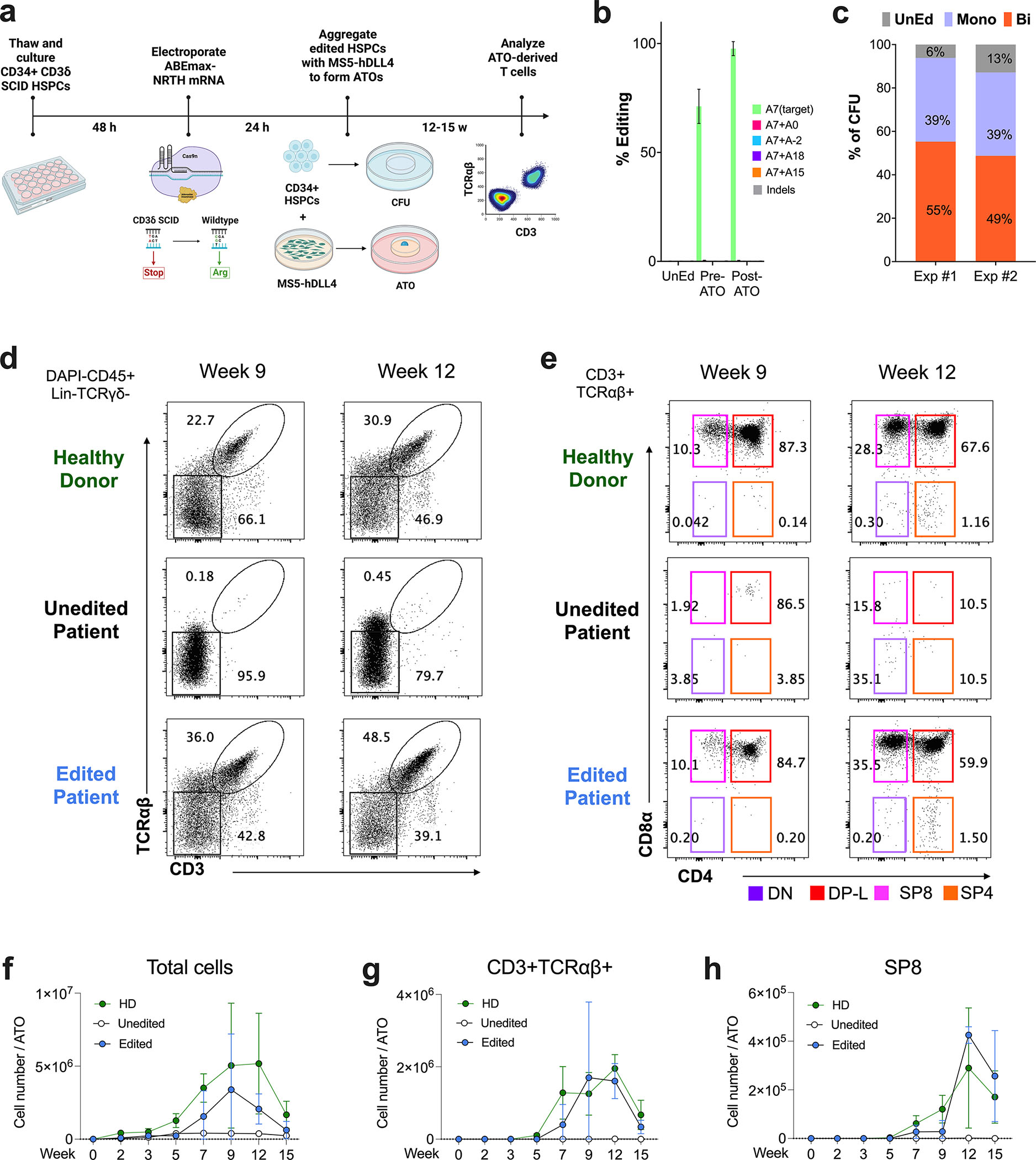
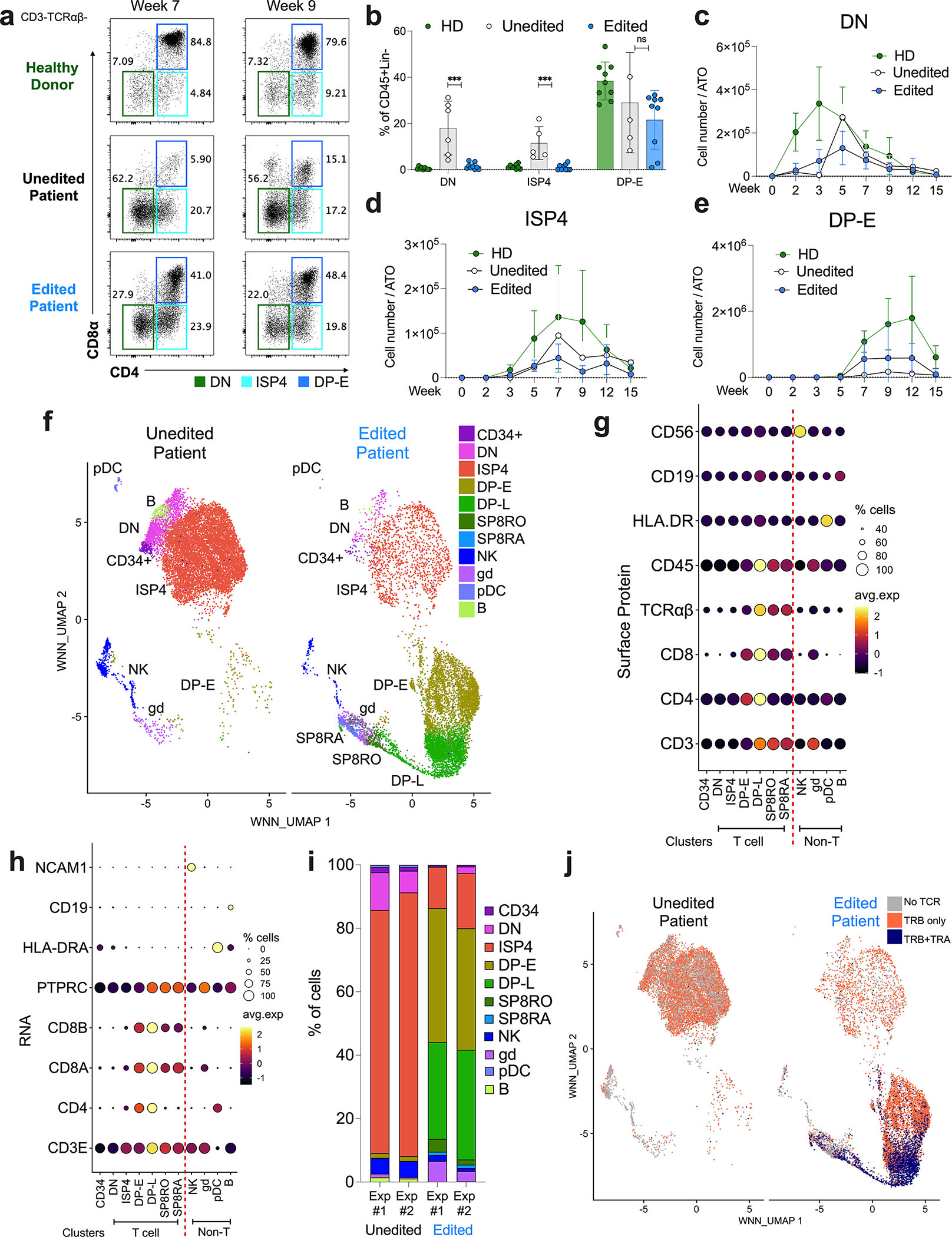
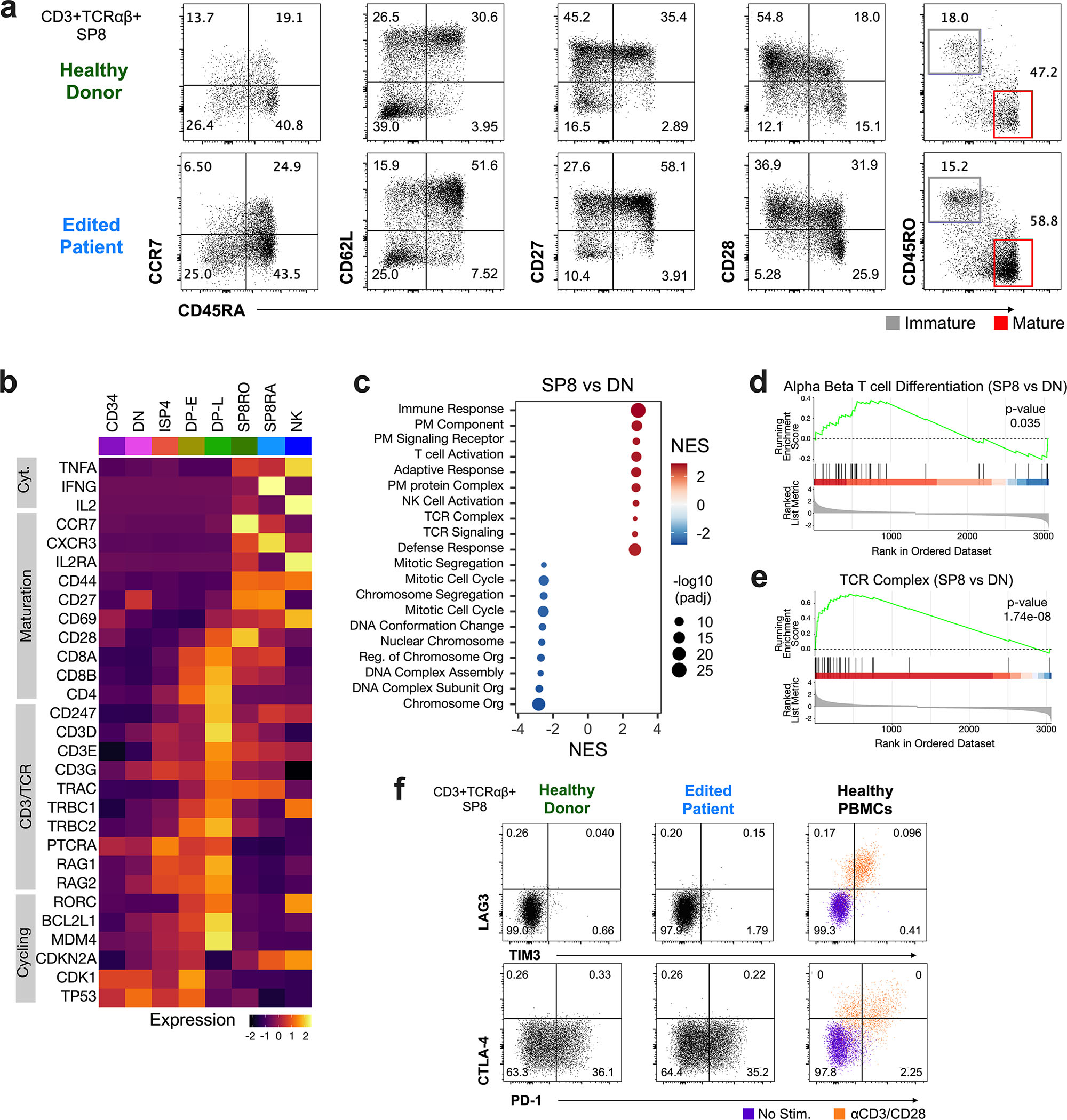
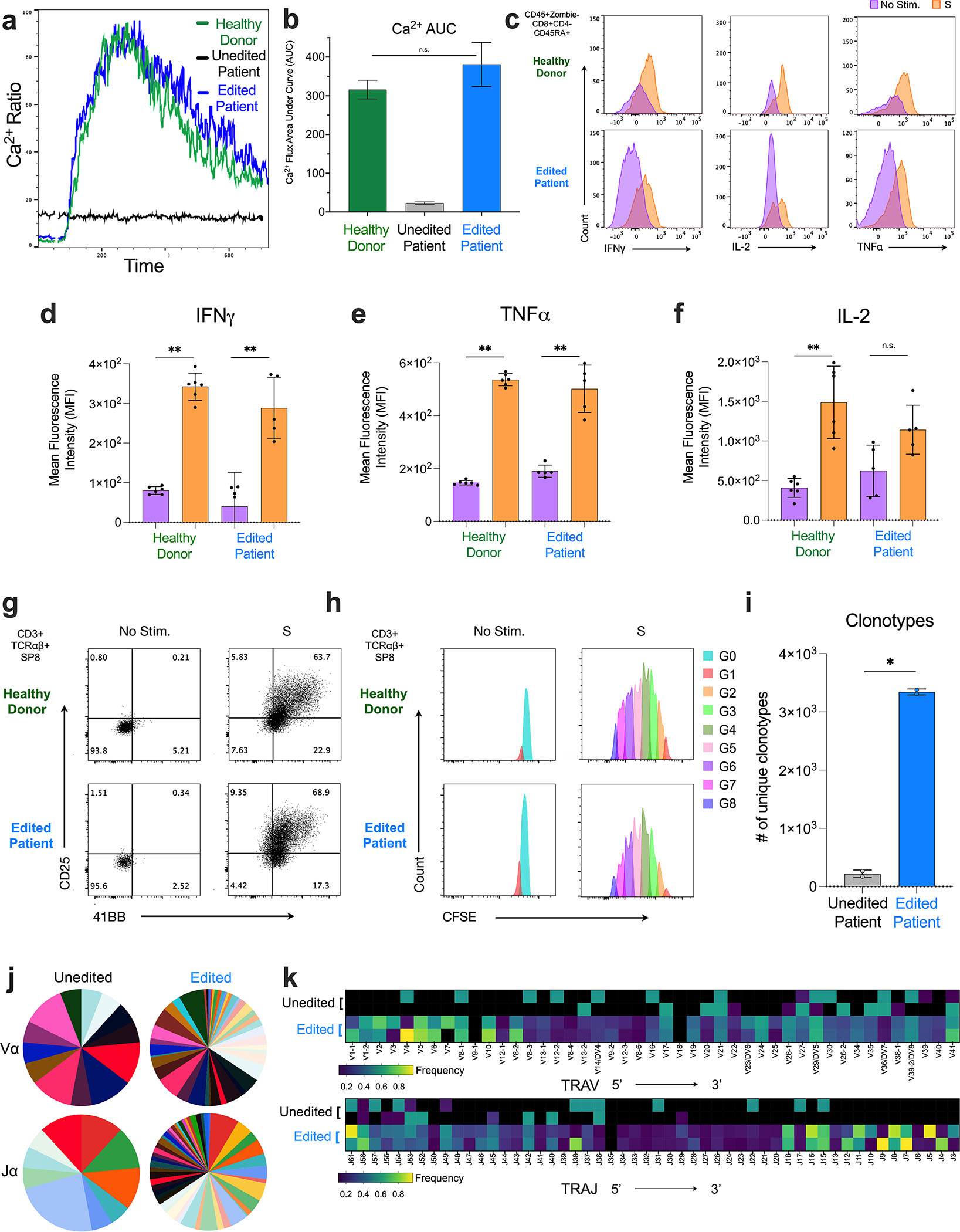
CD3δ SCID is a devastating inborn error of immunity caused by mutations in CD3D, encoding the invariant CD3δ chain of the CD3/TCR complex necessary for normal thymopoiesis. We demonstrate an adenine base editing (ABE) strategy to restore CD3δ in autologous hematopoietic stem and progenitor cells (HSPCs). Delivery of mRNA encoding a laboratory-evolved ABE and guide RNA into a CD3δ SCID patient's HSPCs resulted in a 71.2% ± 7.85% (n = 3) correction of the pathogenic mutation. Edited HSPCs differentiated in artificial thymic organoids produced mature T cells exhibiting diverse TCR repertoires and TCR-dependent functions. Edited human HSPCs transplanted into immunodeficient mice showed 88% reversion of the CD3D defect in human CD34+ cells isolated from mouse bone marrow after 16 weeks, indicating correction of long-term repopulating HSCs. These findings demonstrate the preclinical efficacy of ABE in HSPCs for the treatment of CD3δ SCID, providing a foundation for the development of a one-time treatment for CD3δ SCID patients.
Keywords: CD3D severe combined immune deficiency; CITE-seq; artificial thymic organoid; base editing; hematopoietic stem and progenitor cells.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests G.M.C. and C.S.S. are founders of Pluto Immunotherapeutics Inc. and serve as consultants to this company. D.R.L. is a consultant and equity holder of Prime Medicine, Beam Therapeutics, Pairwise Plants, and Chroma Medicine, companies that use gene editing or genome engineering.
Figures
Comment in
-
Gene therapy for inborn errors of immunity: Base editing comes into play.Cell. 2023 Mar 30;186(7):1302-1304. doi: 10.1016/j.cell.2023.03.001. Cell. 2023. PMID: 37001495 Free PMC article.
-
Base editing: a novel cure for severe combined immunodeficiency.Signal Transduct Target Ther. 2023 Sep 18;8(1):354. doi: 10.1038/s41392-023-01586-2. Signal Transduct Target Ther. 2023. PMID: 37718346 Free PMC article. No abstract available.
References
-
- Garcillán B, Fuentes P, Marin A. v., Megino RF, Chacon-Arguedas D, Mazariegos MS, Jiménez-Reinoso A, Muñoz-Ruiz M, Laborda RG, Cárdenas PP, et al. (2021). CD3G or CD3D Knockdown in Mature, but Not Immature, T Lymphocytes Similarly Cripples the Human TCRαβ Complex. Front Cell Dev Biol 9. 10.3389/fcell.2021.608490. - DOI - PMC - PubMed
-
- Dadi HK, Simon AJ, and Roifman CM (2003). Effect of CD3 d Deficiency on Maturation of a / b and g / d T-Cell Lineages in Severe Combined Immunodeficiency. - PubMed
-
- Marcus N, Takada H, Law J, Cowan MJ, Gil J, Regueiro JR, Plaza Lopez De Sabando D, Lopez-Granados E, Dalal J, Friedrich W, et al. (2011). Hematopoietic stem cell transplantation for CD3δ deficiency. Journal of Allergy and Clinical Immunology 128, 1050–1057. 10.1016/j.jaci.2011.05.031. - DOI - PMC - PubMed
-
- Kohn DB, Booth C, Shaw KL, Xu-Bayford J, Garabedian E, Trevisan V, Carbonaro-Sarracino DA, Soni K, Terrazas D, Snell K, et al. (2021). Autologous Ex Vivo Lentiviral Gene Therapy for Adenosine Deaminase Deficiency. New England Journal of Medicine 384, 2002–2013. 10.1056/nejmoa2027675. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
