

Multi-omics analysis identifies drivers of protein phosphorylation
- PMID: 36944993
- PMCID: PMC10031968
- DOI: 10.1186/s13059-023-02892-2
Multi-omics analysis identifies drivers of protein phosphorylation
Abstract
Background: Phosphorylation of proteins is a key step in the regulation of many cellular processes including activation of enzymes and signaling cascades. The abundance of a phosphorylated peptide (phosphopeptide) is determined by the abundance of its parent protein and the proportion of target sites that are phosphorylated.
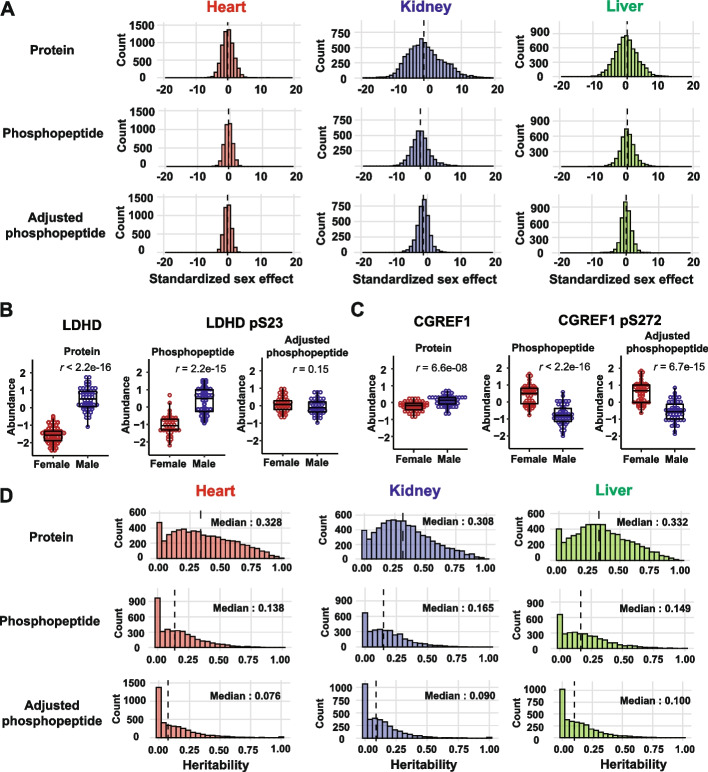
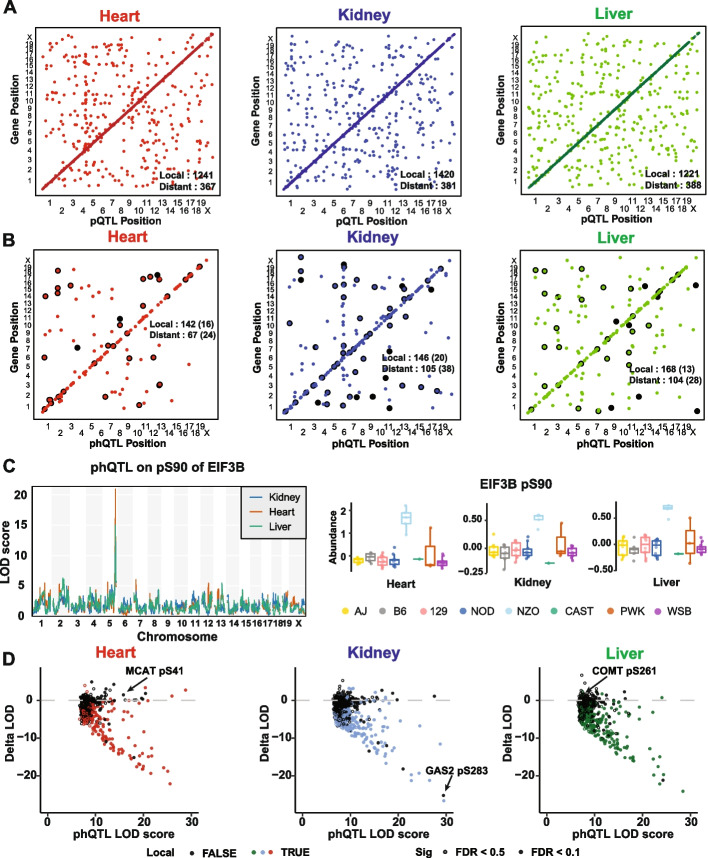
Results: We quantified phosphopeptides, proteins, and transcripts in heart, liver, and kidney tissue samples of mice from 58 strains of the Collaborative Cross strain panel. We mapped ~700 phosphorylation quantitative trait loci (phQTL) across the three tissues and applied genetic mediation analysis to identify causal drivers of phosphorylation. We identified kinases, phosphatases, cytokines, and other factors, including both known and potentially novel interactions between target proteins and genes that regulate site-specific phosphorylation. Our analysis highlights multiple targets of pyruvate dehydrogenase kinase 1 (PDK1), a regulator of mitochondrial function that shows reduced activity in the NZO/HILtJ mouse, a polygenic model of obesity and type 2 diabetes.
Conclusions: Together, this integrative multi-omics analysis in genetically diverse CC strains provides a powerful tool to identify regulators of protein phosphorylation. The data generated in this study provides a resource for further exploration.
Keywords: Collaborative Cross; Medation analysis; Multi-omics; Phosphorylation; Phosphorylation regulation; Quantitative trait loci (QTL).
© 2023. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures





References
-
- Manning G, Whyte DB, Martinez R, Hunter T, Sudarsanam S. The protein kinase complement of the human genome. Science. 1912;2002:298. - PubMed
-
- Haeder M, Rotsch M, Bepler G, Hennig C, Havemann K, Heimann B, Moelling K. Epidermal growth factor receptor expression in human lung cancer cell lines. Cancer Res. 1988;48:1132–1136. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
