

This is a preprint.
Massively parallel characterization of transcriptional regulatory elements in three diverse human cell types
- PMID: 36945371
- PMCID: PMC10028905
- DOI: 10.1101/2023.03.05.531189
Massively parallel characterization of transcriptional regulatory elements in three diverse human cell types
Update in
-
Massively parallel characterization of transcriptional regulatory elements.Nature. 2025 Mar;639(8054):411-420. doi: 10.1038/s41586-024-08430-9. Epub 2025 Jan 15. Nature. 2025. PMID: 39814889 Free PMC article.
Abstract
The human genome contains millions of candidate cis-regulatory elements (CREs) with cell-type-specific activities that shape both health and myriad disease states. However, we lack a functional understanding of the sequence features that control the activity and cell-type-specific features of these CREs. Here, we used lentivirus-based massively parallel reporter assays (lentiMPRAs) to test the regulatory activity of over 680,000 sequences, representing a nearly comprehensive set of all annotated CREs among three cell types (HepG2, K562, and WTC11), finding 41.7% to be functional. By testing sequences in both orientations, we find promoters to have significant strand orientation effects. We also observe that their 200 nucleotide cores function as non-cell-type-specific 'on switches' providing similar expression levels to their associated gene. In contrast, enhancers have weaker orientation effects, but increased tissue-specific characteristics. Utilizing our lentiMPRA data, we develop sequence-based models to predict CRE function with high accuracy and delineate regulatory motifs. Testing an additional lentiMPRA library encompassing 60,000 CREs in all three cell types, we further identified factors that determine cell-type specificity. Collectively, our work provides an exhaustive catalog of functional CREs in three widely used cell lines, and showcases how large-scale functional measurements can be used to dissect regulatory grammar.
Conflict of interest statement
Competing interests. V.A. is an employee of Sanofi Pasteur Inc. J.S. is a scientific advisory board member, consultant and/or co-founder of Cajal Neuroscience, Guardant Health, Maze Therapeutics, Camp4 Therapeutics, Phase Genomics, Adaptive Biotechnologies, Scale Biosciences, Sixth Street Capital, and Pacific Biosciences. N.A. is the cofounder and on the scientific advisory board of Regel Therapeutics and receives funding from BioMarin Pharmaceutical Incorporated. All other authors declare no competing interests.
Figures








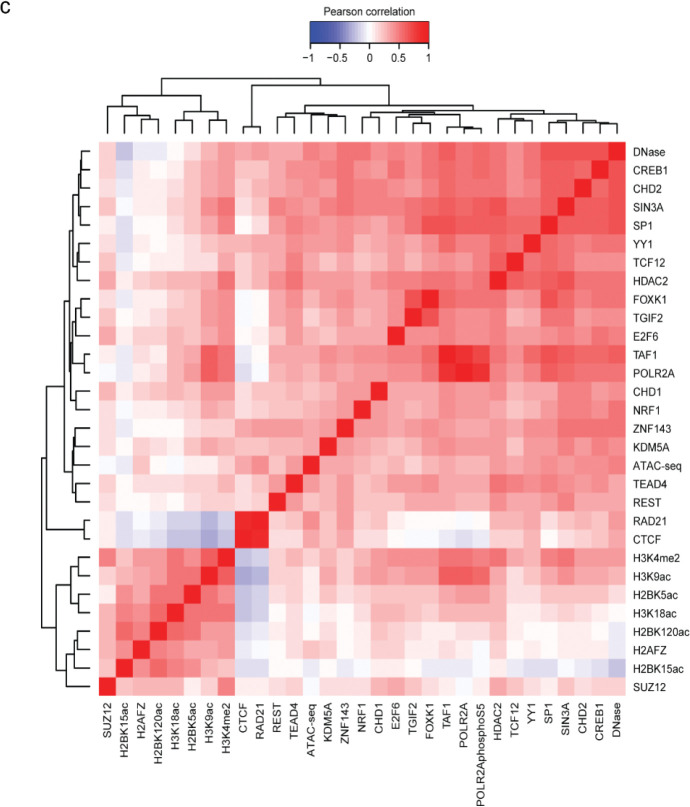
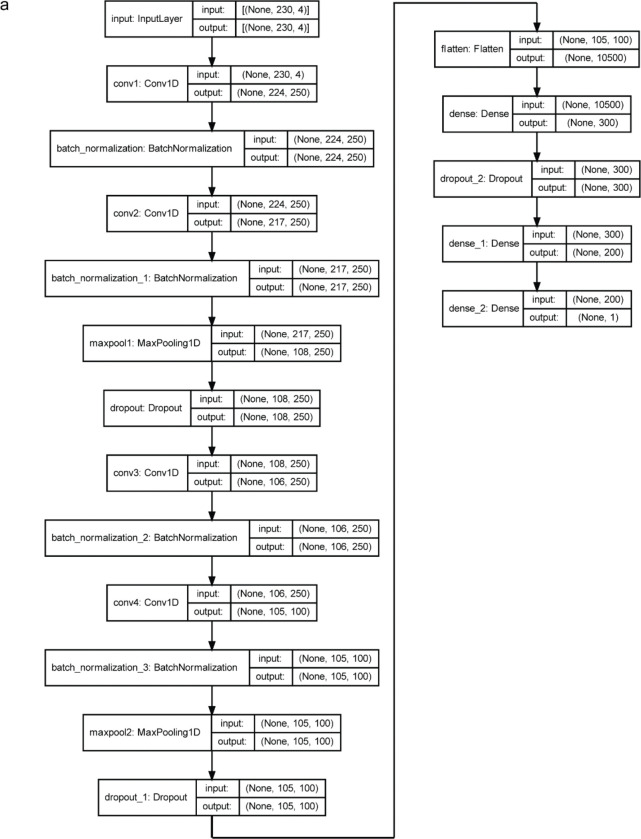





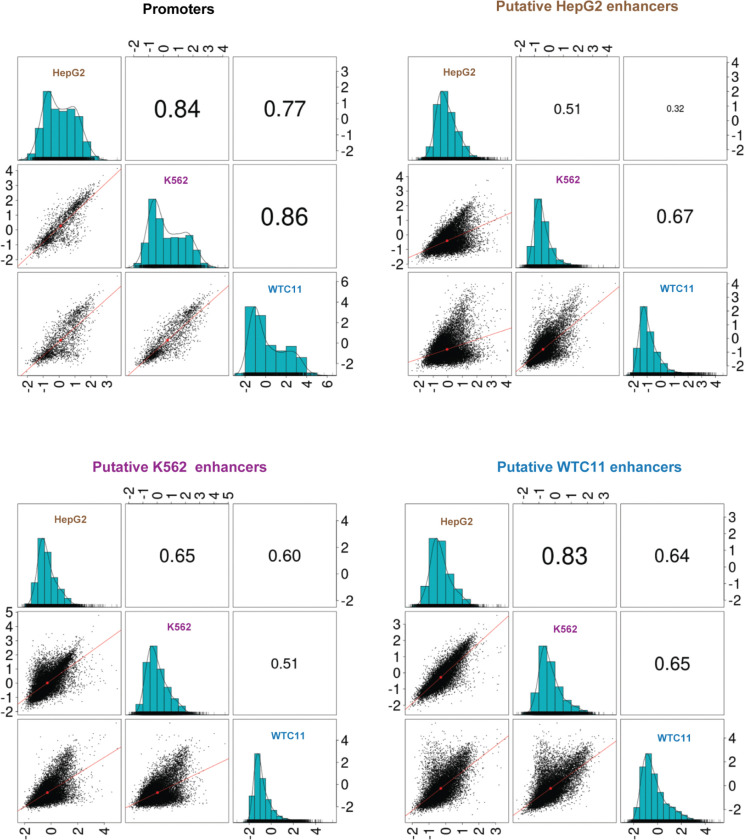




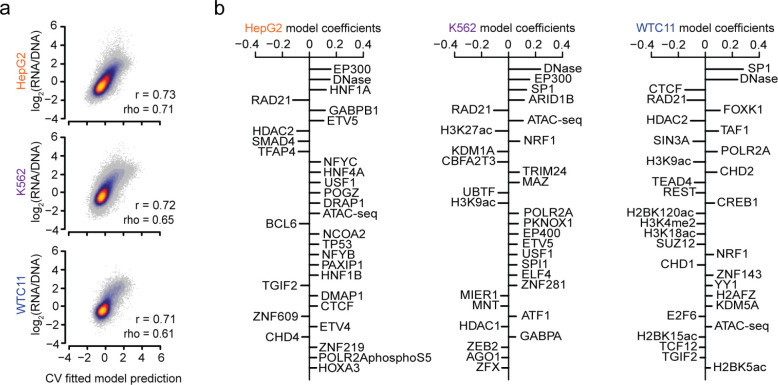


References
-
- Rickels R. & Shilatifard A. Enhancer Logic and Mechanics in Development and Disease. Trends Cell Biol. 28, 608–630 (2018). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials