

This is a preprint.
RNA Polymerase II pausing temporally coordinates cell cycle progression and erythroid differentiation
- PMID: 36945604
- PMCID: PMC10029049
- DOI: 10.1101/2023.03.03.23286760
RNA Polymerase II pausing temporally coordinates cell cycle progression and erythroid differentiation
Update in
-
RNA polymerase II pausing temporally coordinates cell cycle progression and erythroid differentiation.Dev Cell. 2023 Oct 23;58(20):2112-2127.e4. doi: 10.1016/j.devcel.2023.07.018. Epub 2023 Aug 15. Dev Cell. 2023. PMID: 37586368 Free PMC article.
Abstract
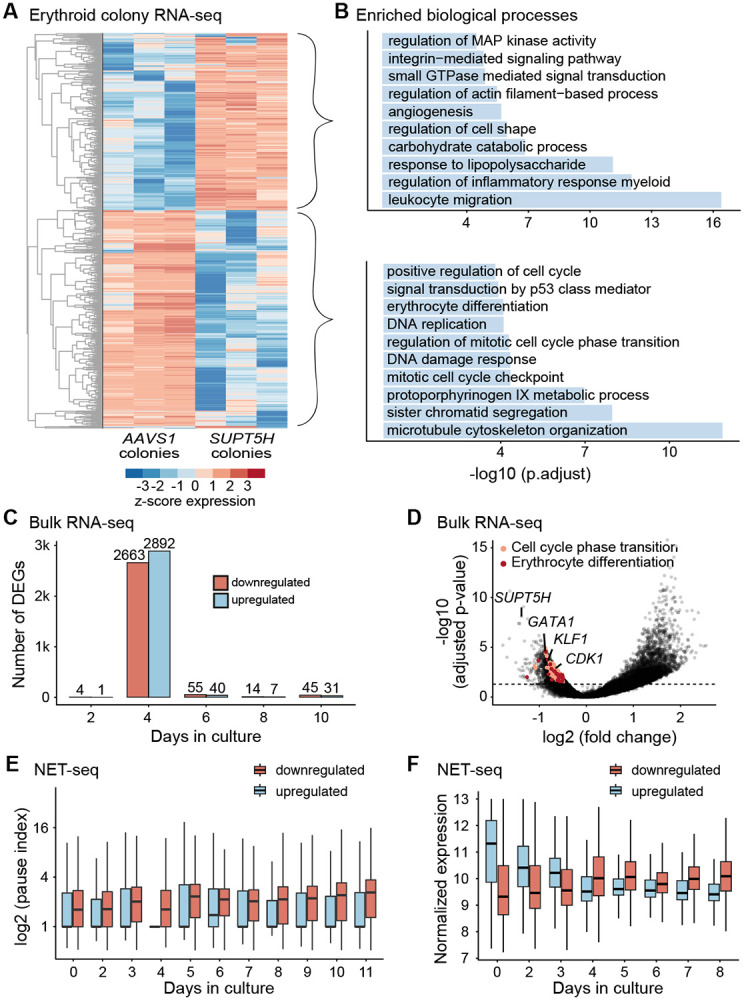
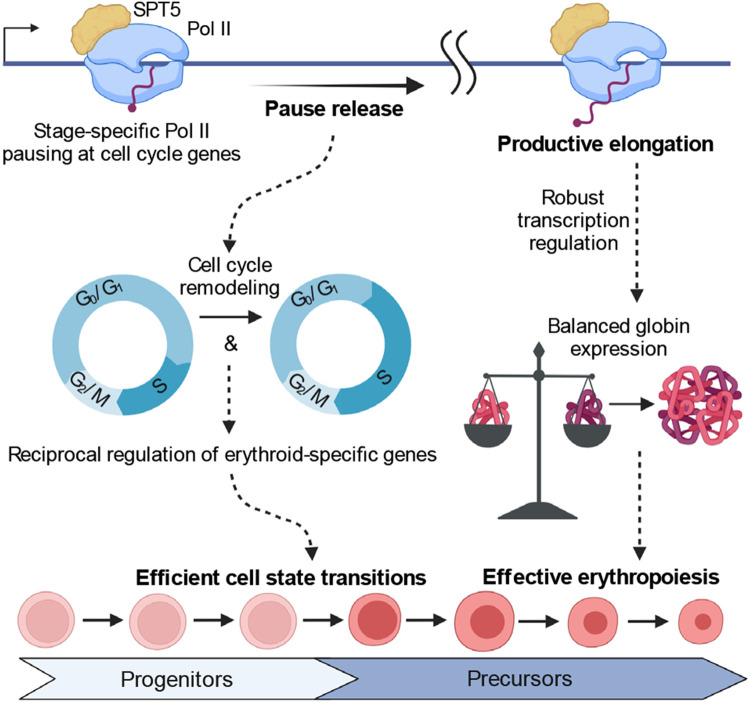
The controlled release of promoter-proximal paused RNA polymerase II (Pol II) into productive elongation is a major step in gene regulation. However, functional analysis of Pol II pausing is difficult because factors that regulate pause release are almost all essential. In this study, we identified heterozygous loss-of-function mutations in SUPT5H , which encodes SPT5, in individuals with β-thalassemia unlinked to HBB mutations. During erythropoiesis in healthy human cells, cell cycle genes were highly paused at the transition from progenitors to precursors. When the pathogenic mutations were recapitulated by SUPT5H editing, Pol II pause release was globally disrupted, and the transition from progenitors to precursors was delayed, marked by a transient lag in erythroid-specific gene expression and cell cycle kinetics. Despite this delay, cells terminally differentiate, and cell cycle phase distributions normalize. Therefore, hindering pause release perturbs proliferation and differentiation dynamics at a key transition during erythropoiesis, revealing a role for Pol II pausing in the temporal coordination between the cell cycle and differentiation.
Conflict of interest statement
Declaration of interests
V.G.S. serves as an advisor to and/or has equity in Branch Biosciences, Ensoma, Novartis, Forma, Sana Biotechnology, and Cellarity, all unrelated to the present work. J.C.U. is an employee of Illumina, Inc., unrelated to the present work. R.I. is a founder, board member and shareholder of Cellforma, unrelated to the present work.
Figures





References
-
- Abuhashem Abderhman, Chivu Alexandra G., Zhao Yixin, Rice Edward J., Siepel Adam, Danko Charles G., and Hadjantonakis Anna-Katerina. 2022. “RNA Pol II Pausing Facilitates Phased Pluripotency Transitions by Buffering Transcription.” Genes & Development, August. 10.1101/gad.349565.122. - DOI - PMC - PubMed
-
- Achour Ahlem, Koopmann Tamara T., Castel Rob, Santen Gijs W. E., Hollander den Nicolette, Knijnenburg Jeroen, Ruivenkamp Claudia A. L., et al. 2020. “A New Gene Associated with a Beta-Thalassemia Phenotype: The Observation of Variants in SUPT5H.” Blood, June. 10.1182/blood.2020005934. - DOI - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources