

Ultrafast structural changes direct the first molecular events of vision
- PMID: 36949205
- PMCID: PMC10060157
- DOI: 10.1038/s41586-023-05863-6
Ultrafast structural changes direct the first molecular events of vision
Abstract
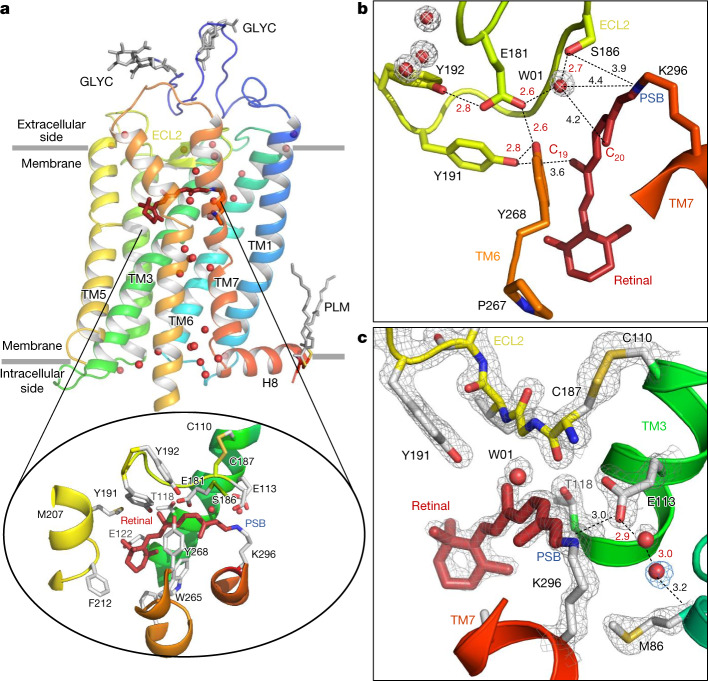
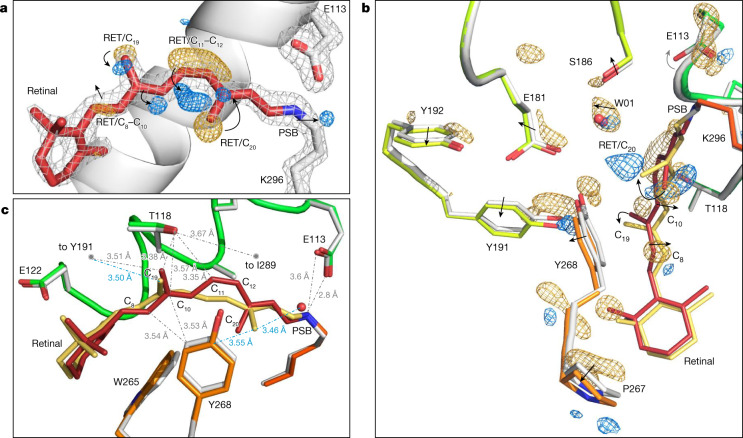
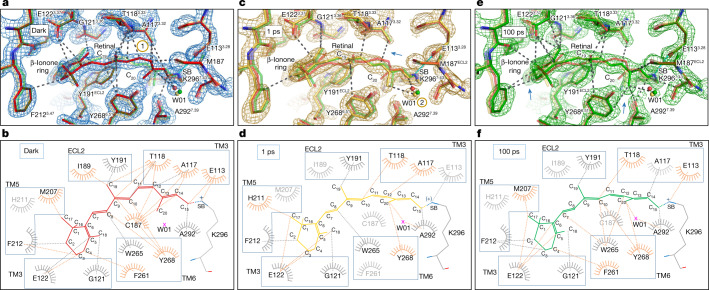
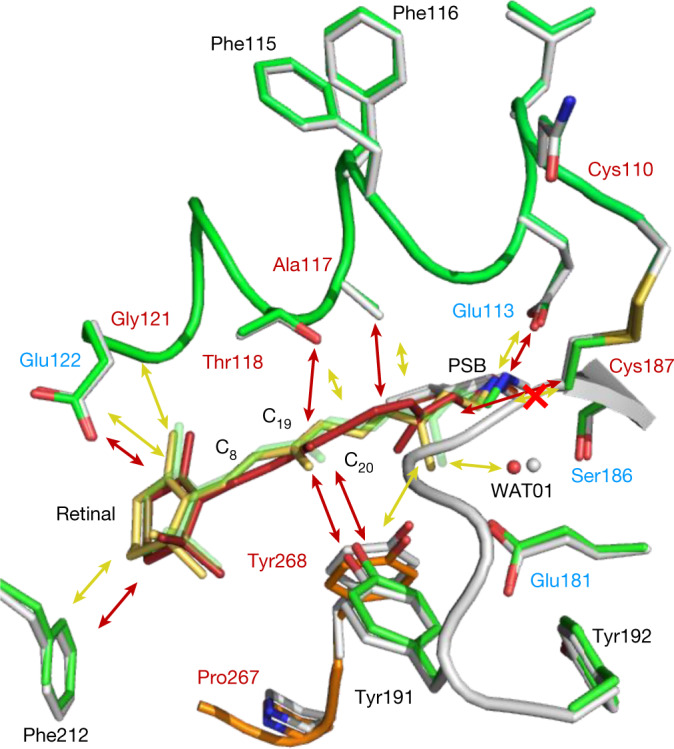
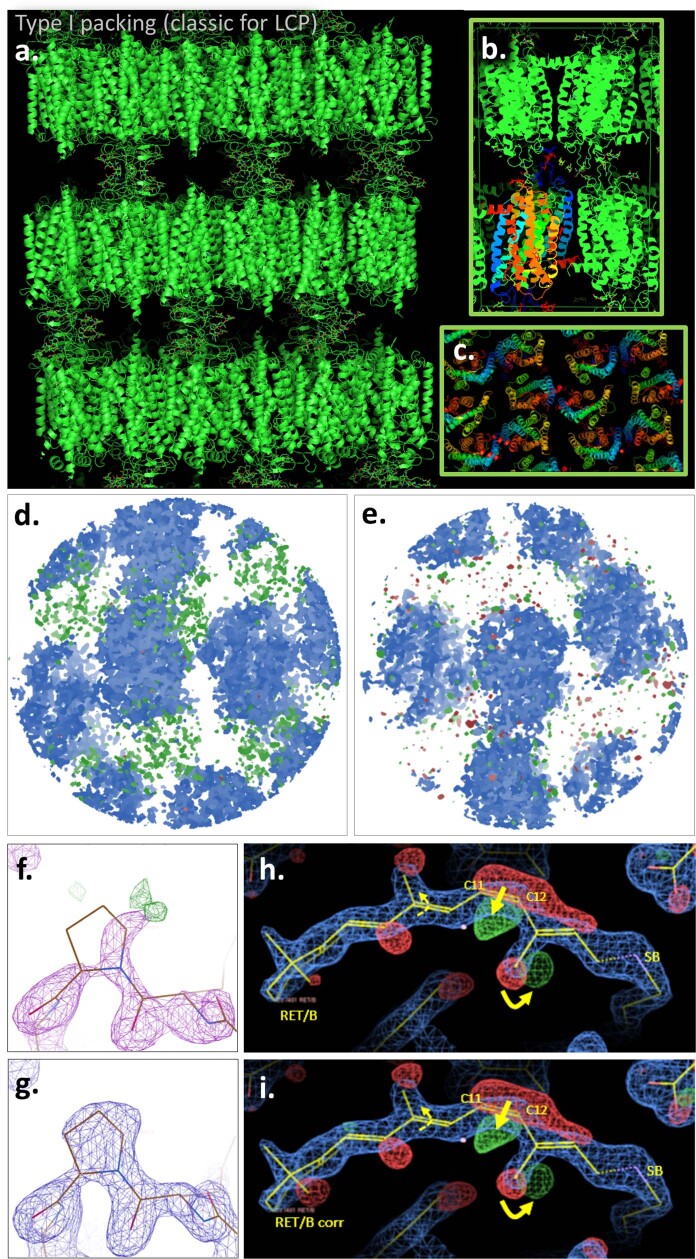
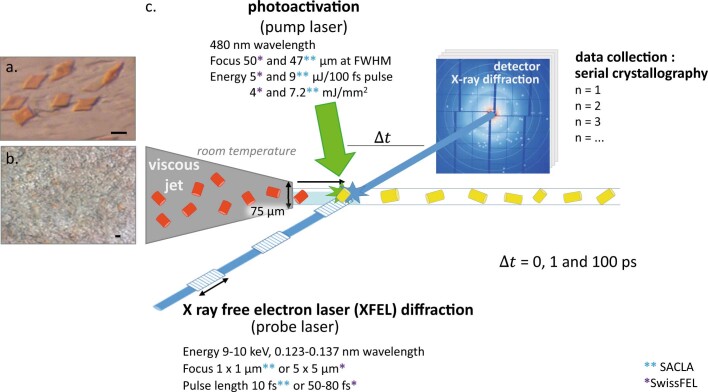
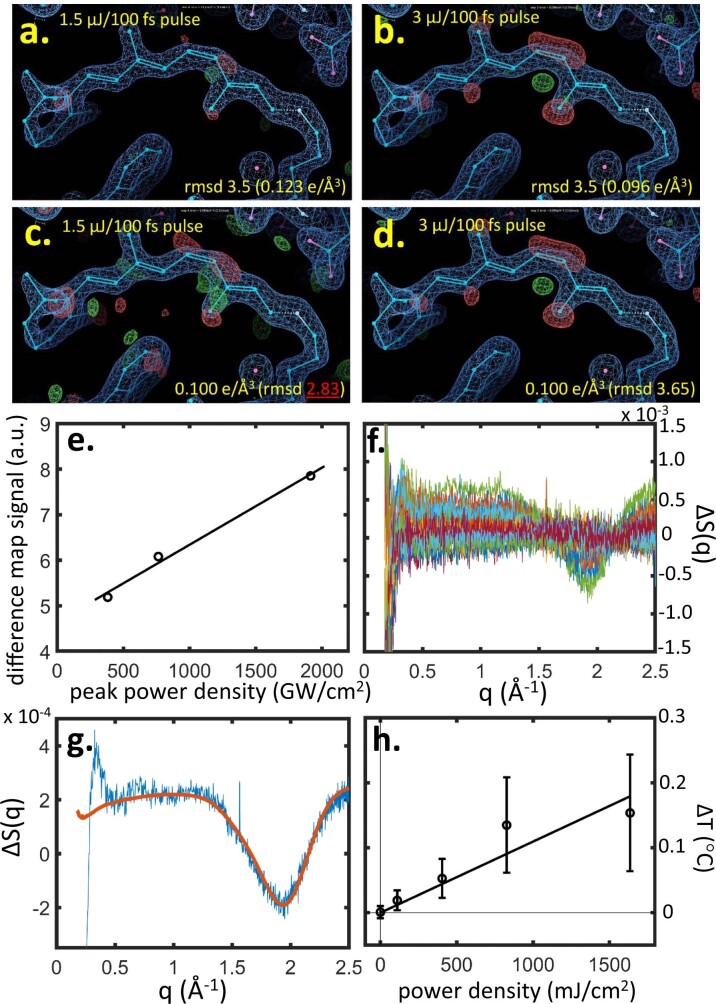
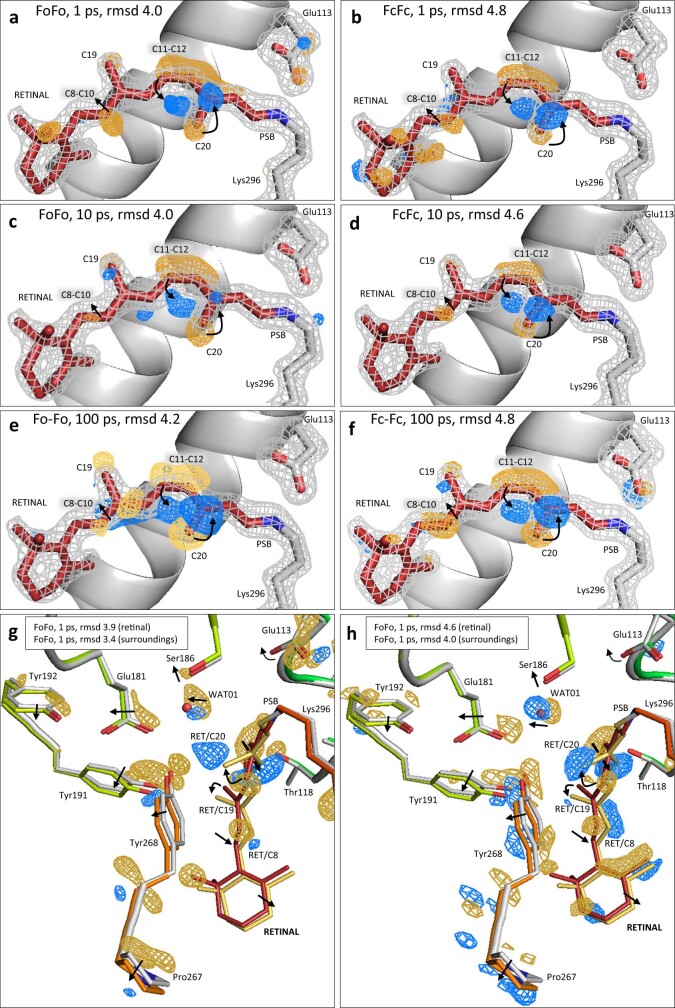
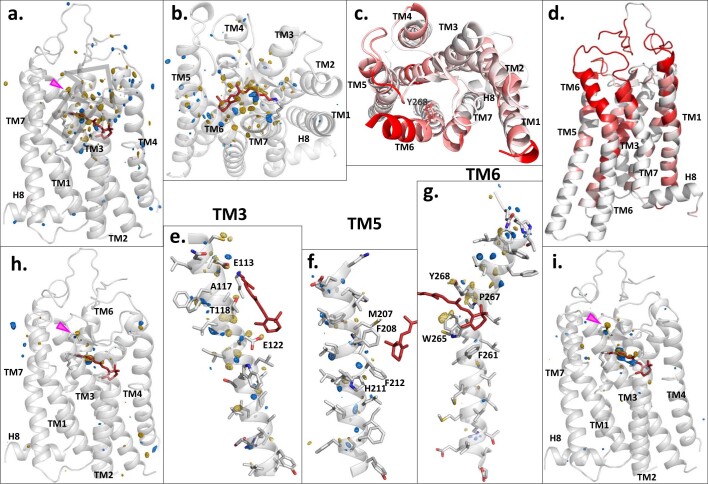
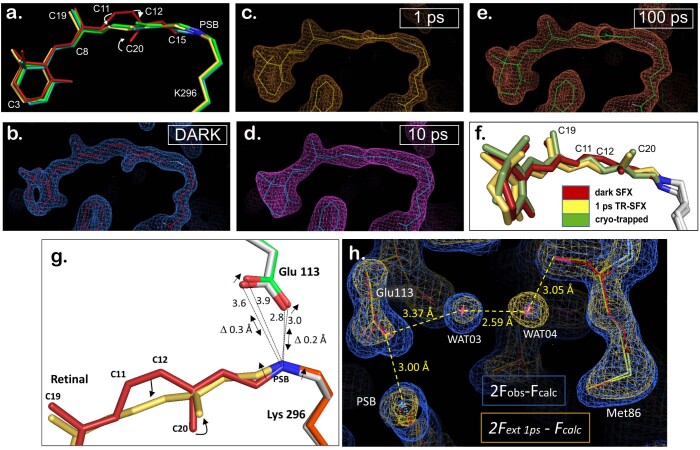
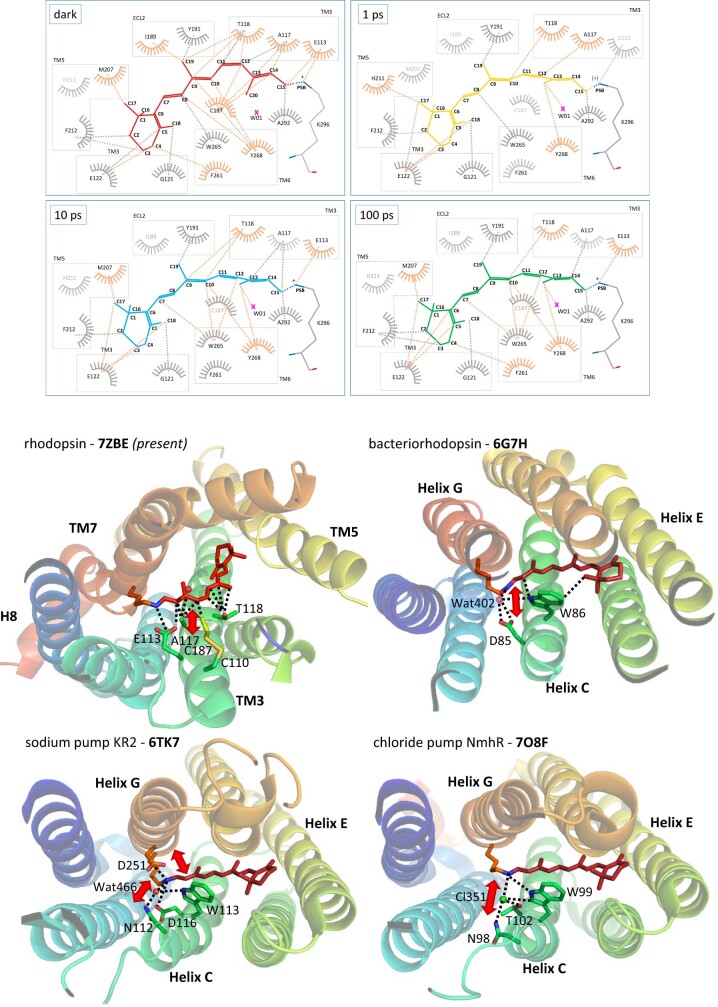
Vision is initiated by the rhodopsin family of light-sensitive G protein-coupled receptors (GPCRs)1. A photon is absorbed by the 11-cis retinal chromophore of rhodopsin, which isomerizes within 200 femtoseconds to the all-trans conformation2, thereby initiating the cellular signal transduction processes that ultimately lead to vision. However, the intramolecular mechanism by which the photoactivated retinal induces the activation events inside rhodopsin remains experimentally unclear. Here we use ultrafast time-resolved crystallography at room temperature3 to determine how an isomerized twisted all-trans retinal stores the photon energy that is required to initiate the protein conformational changes associated with the formation of the G protein-binding signalling state. The distorted retinal at a 1-ps time delay after photoactivation has pulled away from half of its numerous interactions with its binding pocket, and the excess of the photon energy is released through an anisotropic protein breathing motion in the direction of the extracellular space. Notably, the very early structural motions in the protein side chains of rhodopsin appear in regions that are involved in later stages of the conserved class A GPCR activation mechanism. Our study sheds light on the earliest stages of vision in vertebrates and points to fundamental aspects of the molecular mechanisms of agonist-mediated GPCR activation.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
Earliest molecular events of vision revealed.Nature. 2023 Mar;615(7954):802-803. doi: 10.1038/d41586-023-00504-4. Nature. 2023. PMID: 36949122 No abstract available.
