

Memory T cells possess an innate-like function in local protection from mucosal infection
- PMID: 36951943
- PMCID: PMC10178838
- DOI: 10.1172/JCI162800
Memory T cells possess an innate-like function in local protection from mucosal infection
Abstract
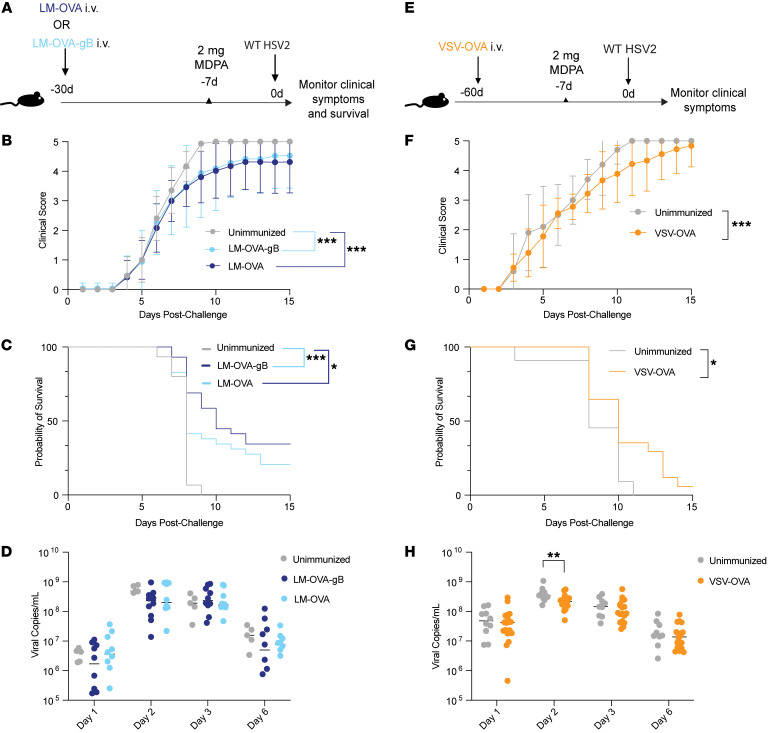
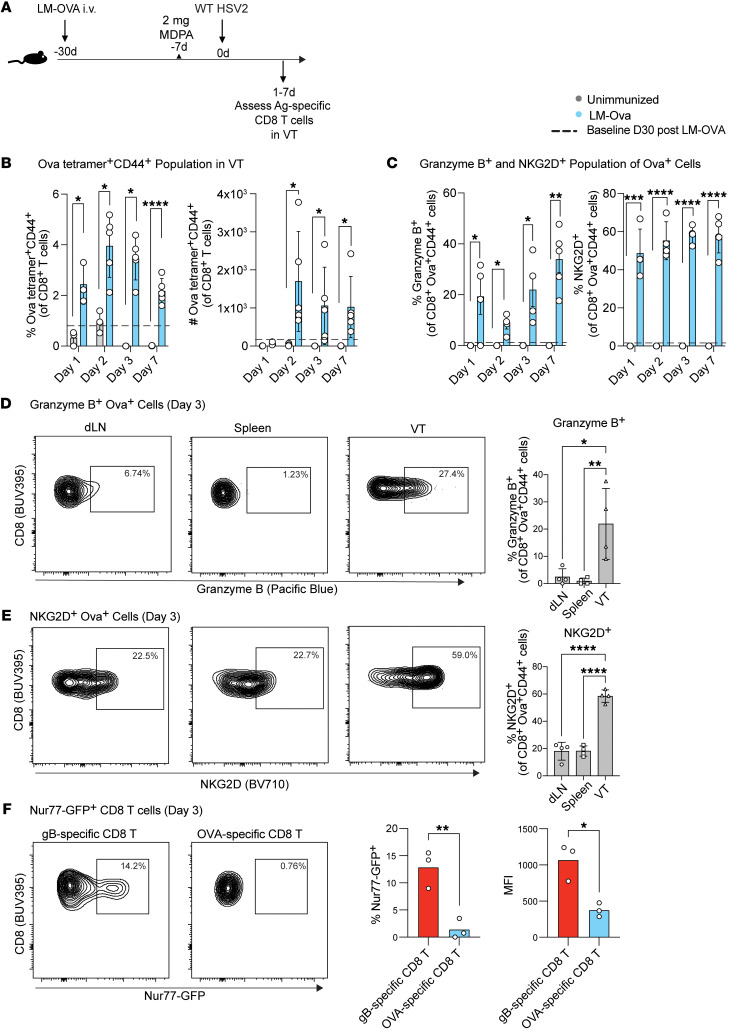
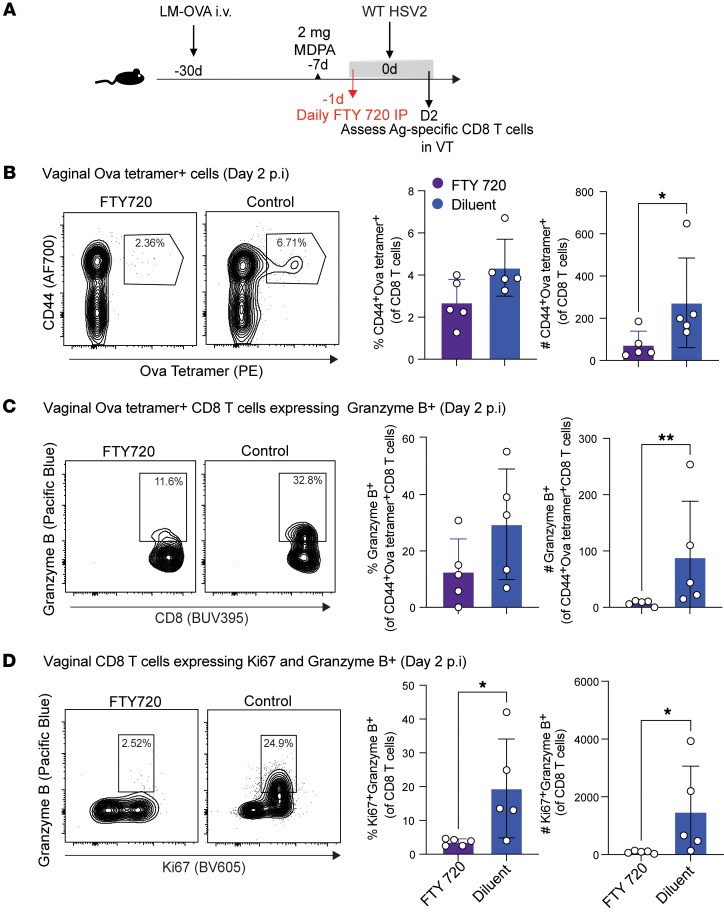
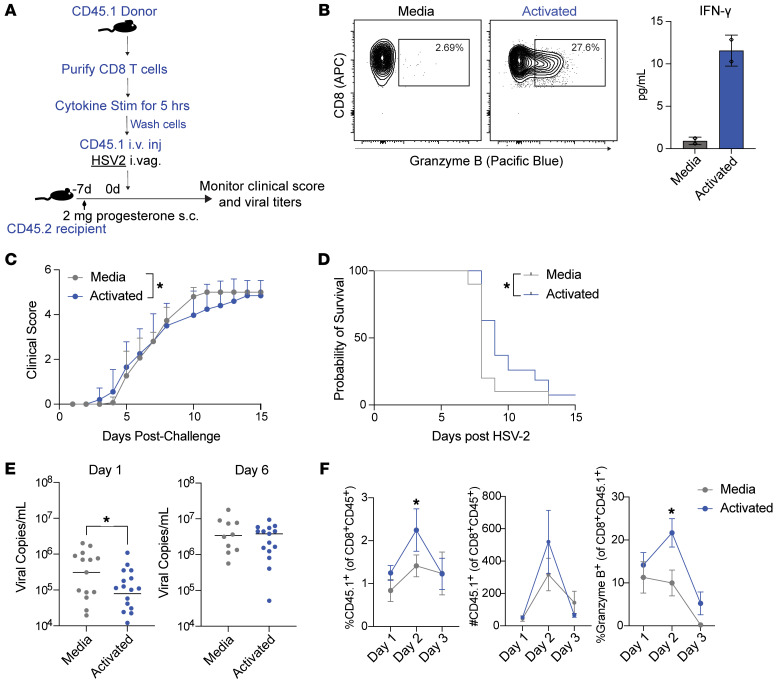
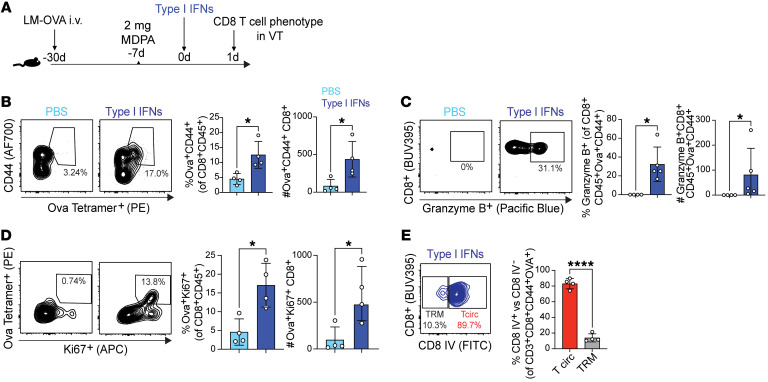
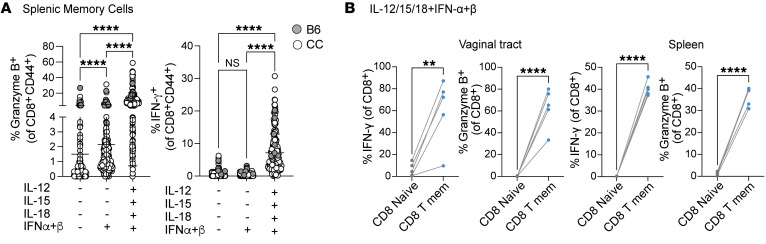
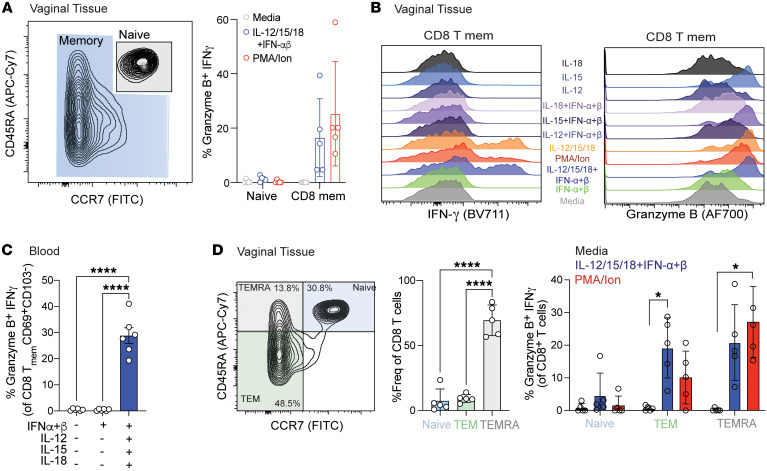
Mucosal infections pose a significant global health burden. Antigen-specific tissue-resident T cells are critical to maintaining barrier immunity. Previous studies in the context of systemic infection suggest that memory CD8+ T cells may also provide innate-like protection against antigenically unrelated pathogens independent of T cell receptor engagement. Whether bystander T cell activation is also an important defense mechanism in the mucosa is poorly understood. Here, we investigated whether innate-like memory CD8+ T cells could protect against a model mucosal virus infection, herpes simplex virus 2 (HSV-2). We found that immunization with an irrelevant antigen delayed disease progression from lethal HSV-2 challenge, suggesting that memory CD8+ T cells may mediate protection despite the lack of antigen specificity. Upon HSV-2 infection, we observed an early infiltration, rather than substantial local proliferation, of antigen-nonspecific CD8+ T cells, which became bystander-activated only within the infected mucosal tissue. Critically, we show that bystander-activated CD8+ T cells are sufficient to reduce early viral burden after HSV-2 infection. Finally, local cytokine cues within the tissue microenvironment after infection were sufficient for bystander activation of mucosal tissue memory CD8+ T cells from mice and humans. Altogether, our findings suggest that local bystander activation of CD8+ memory T cells contributes a fast and effective innate-like response to infection in mucosal tissue.
Keywords: Immunology; Infectious disease; T cells.
Conflict of interest statement
Figures
Similar articles
-
CXCL10/CXCR3-Dependent Mobilization of Herpes Simplex Virus-Specific CD8+ TEM and CD8+ TRM Cells within Infected Tissues Allows Efficient Protection against Recurrent Herpesvirus Infection and Disease.J Virol. 2017 Jun 26;91(14):e00278-17. doi: 10.1128/JVI.00278-17. Print 2017 Jul 15. J Virol. 2017. PMID: 28468883 Free PMC article.
-
Estradiol Enhances Antiviral CD4+ Tissue-Resident Memory T Cell Responses following Mucosal Herpes Simplex Virus 2 Vaccination through an IL-17-Mediated Pathway.J Virol. 2020 Dec 9;95(1):e01206-20. doi: 10.1128/JVI.01206-20. Print 2020 Dec 9. J Virol. 2020. PMID: 33028712 Free PMC article.
-
Human Asymptomatic Epitopes Identified from the Herpes Simplex Virus Tegument Protein VP13/14 (UL47) Preferentially Recall Polyfunctional Effector Memory CD44high CD62Llow CD8+ TEM Cells and Protect Humanized HLA-A*02:01 Transgenic Mice against Ocular Herpesvirus Infection.J Virol. 2017 Jan 3;91(2):e01793-16. doi: 10.1128/JVI.01793-16. Print 2017 Jan 15. J Virol. 2017. PMID: 27847359 Free PMC article.
-
Bystander activation in memory and antigen-inexperienced memory-like CD8 T cells.Curr Opin Immunol. 2023 Jun;82:102299. doi: 10.1016/j.coi.2023.102299. Epub 2023 Mar 11. Curr Opin Immunol. 2023. PMID: 36913776 Review.
-
Immunological control of herpes simplex virus infections.J Neurovirol. 2013 Aug;19(4):328-45. doi: 10.1007/s13365-013-0189-3. Epub 2013 Aug 14. J Neurovirol. 2013. PMID: 23943467 Free PMC article. Review.
Cited by
-
The Epithelial Immune Response to Human Papillomavirus Infection.Pathogens. 2025 May 9;14(5):464. doi: 10.3390/pathogens14050464. Pathogens. 2025. PMID: 40430784 Free PMC article. Review.
-
Steady-state, therapeutic, and helminth-induced IL-4 compromise protective CD8 T cell bystander activation.bioRxiv [Preprint]. 2024 Jun 12:2024.06.10.598293. doi: 10.1101/2024.06.10.598293. bioRxiv. 2024. PMID: 38915668 Free PMC article. Preprint.
-
IL-15-Activated CD38+HLA-DR+CD8+ T cells induce liver injury in cirrhosis via JAK/STAT5 and PI3K/mTOR pathways.Sci Rep. 2025 May 21;15(1):17612. doi: 10.1038/s41598-025-02693-6. Sci Rep. 2025. PMID: 40399368 Free PMC article.
-
Pivotal role of tissue-resident memory lymphocytes in the control of mucosal infections: can mucosal vaccination induce protective tissue-resident memory T and B cells?Front Immunol. 2023 Sep 11;14:1216402. doi: 10.3389/fimmu.2023.1216402. eCollection 2023. Front Immunol. 2023. PMID: 37753095 Free PMC article. No abstract available.
-
Recurrent infection transiently expands human tissue T cells while maintaining long-term homeostasis.J Exp Med. 2023 Sep 4;220(9):e20210692. doi: 10.1084/jem.20210692. Epub 2023 Jun 14. J Exp Med. 2023. PMID: 37314481 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
