

A gut-secreted peptide suppresses arousability from sleep
- PMID: 36958331
- PMCID: PMC10216829
- DOI: 10.1016/j.cell.2023.02.022
A gut-secreted peptide suppresses arousability from sleep
Erratum in
-
A gut-secreted peptide suppresses arousability from sleep.Cell. 2023 May 11;186(10):2273-2274. doi: 10.1016/j.cell.2023.04.005. Cell. 2023. PMID: 37172567 Free PMC article. No abstract available.
Abstract
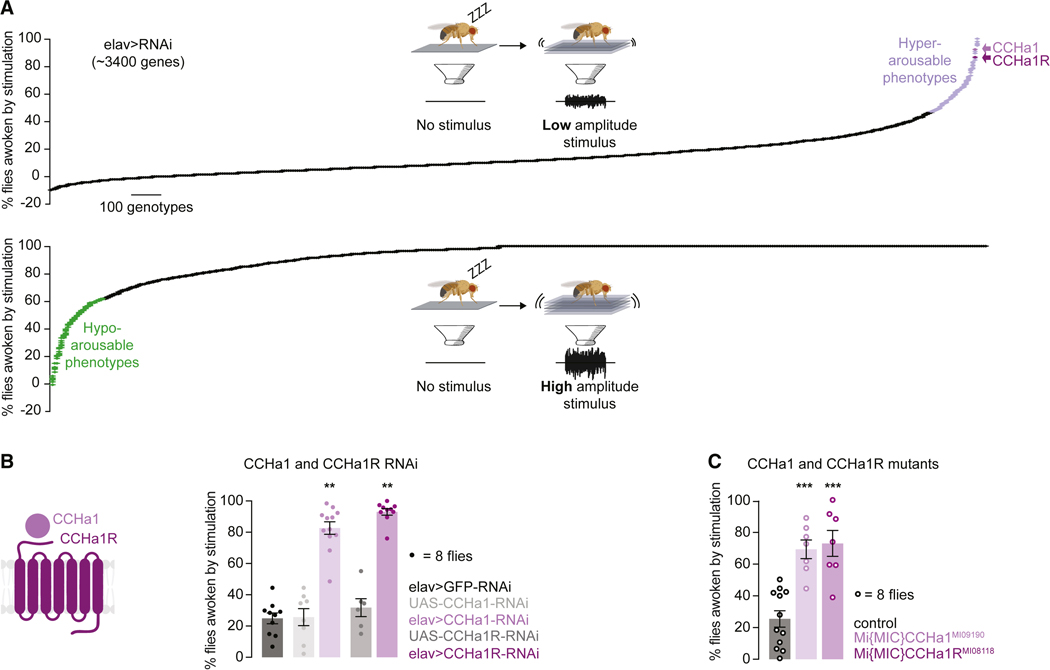
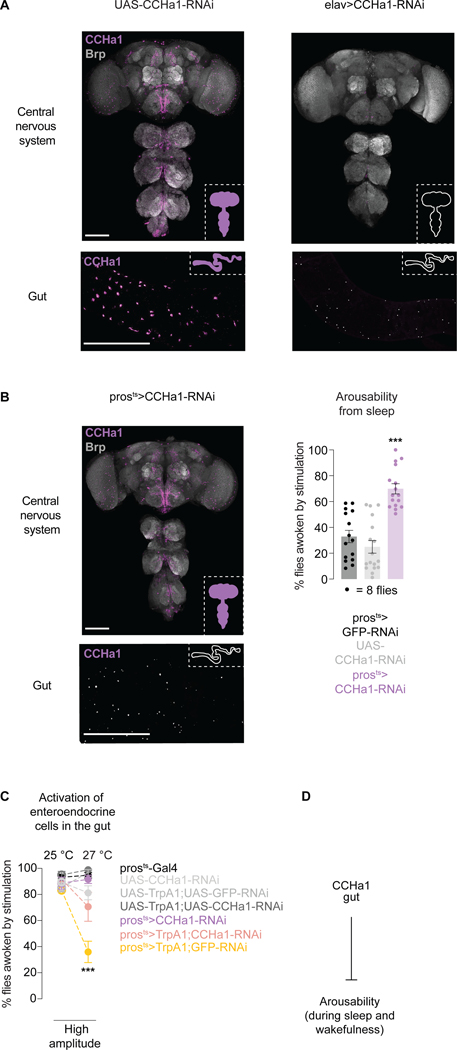
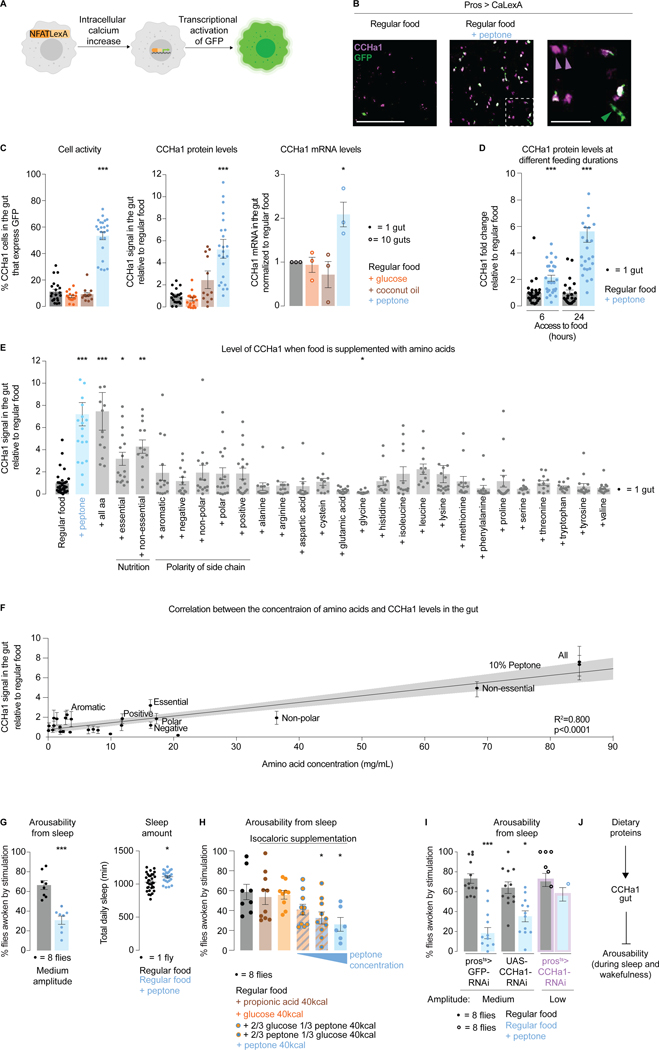
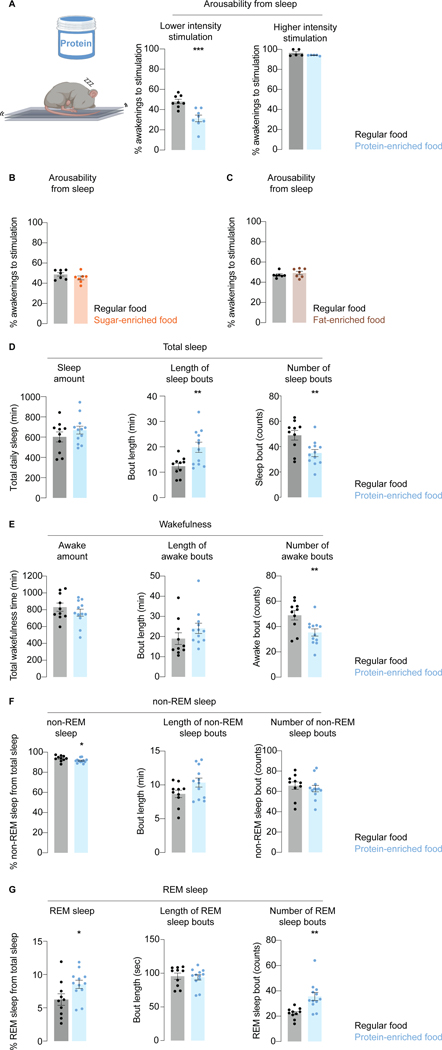
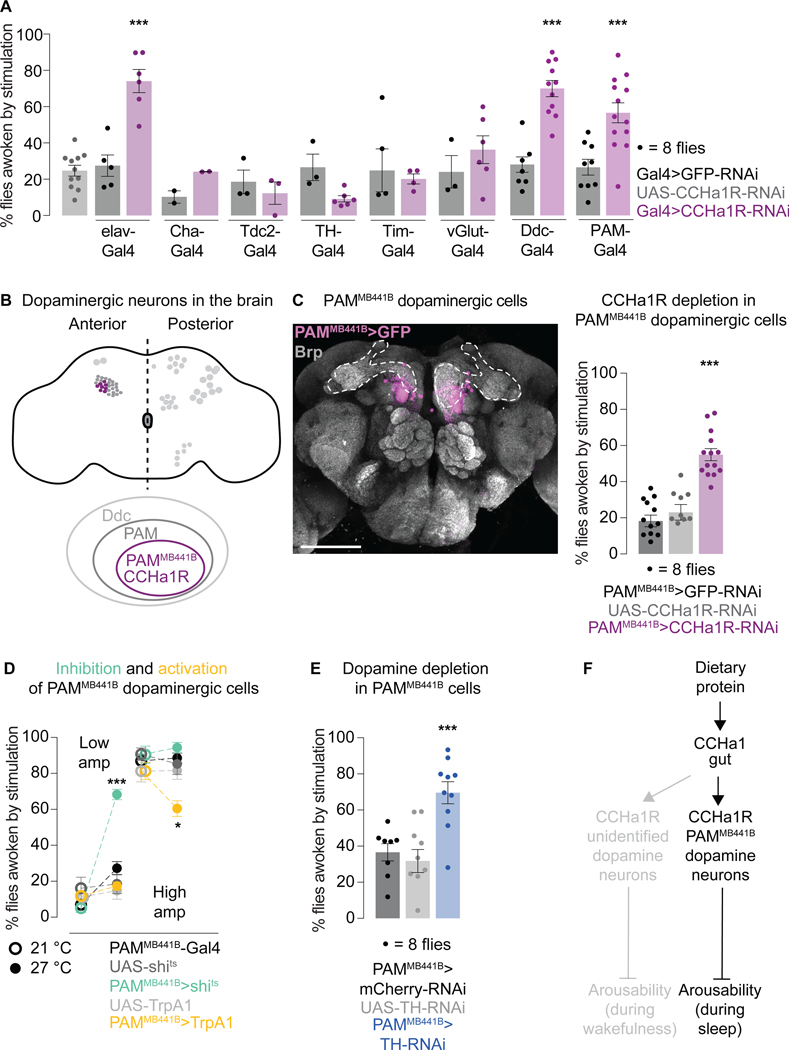
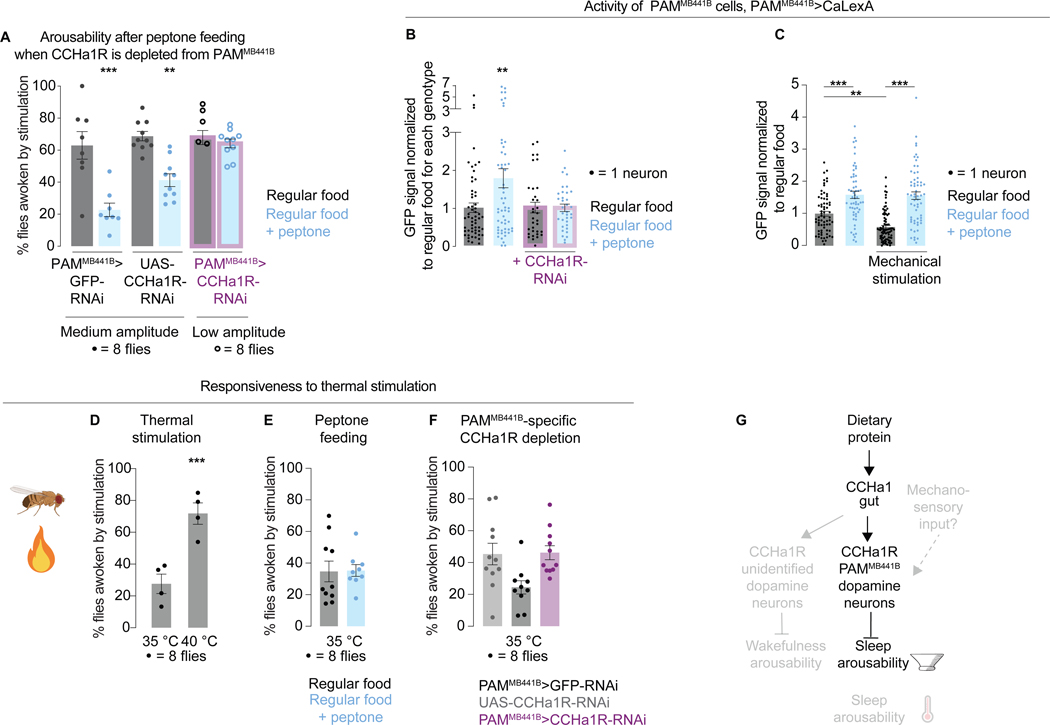
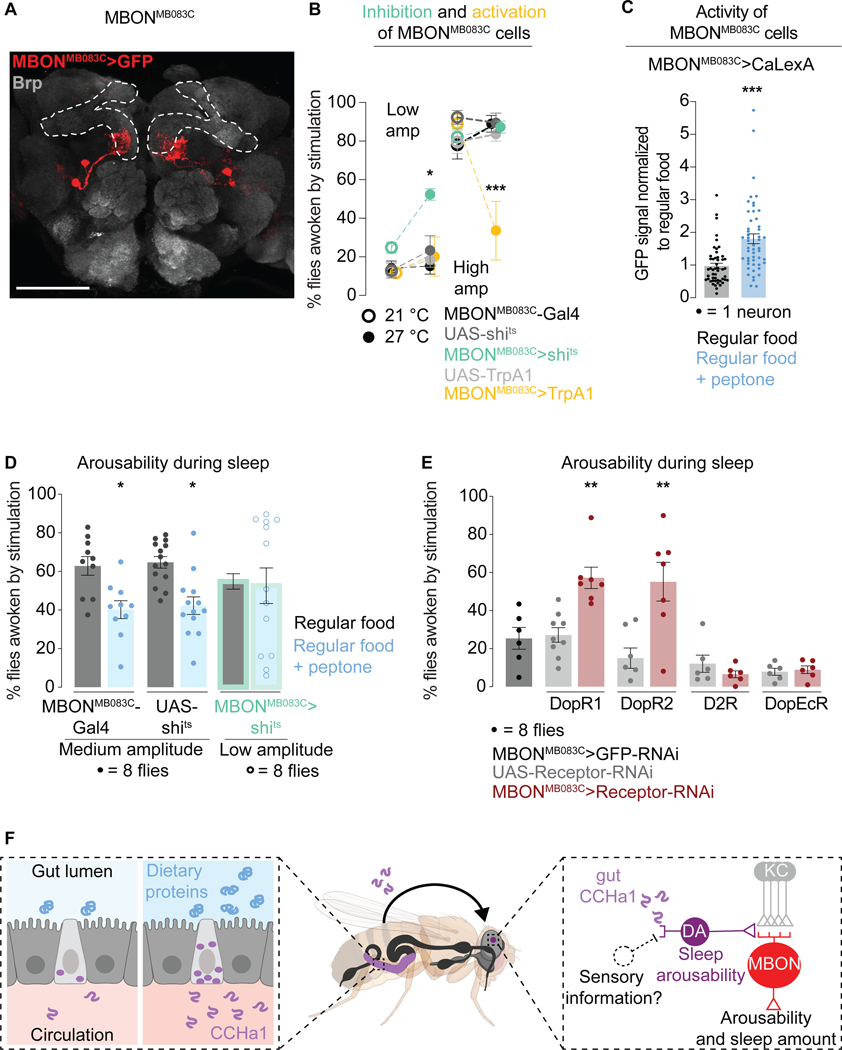
Suppressing sensory arousal is critical for sleep, with deeper sleep requiring stronger sensory suppression. The mechanisms that enable sleeping animals to largely ignore their surroundings are not well understood. We show that the responsiveness of sleeping flies and mice to mechanical vibrations is better suppressed when the diet is protein rich. In flies, we describe a signaling pathway through which information about ingested proteins is conveyed from the gut to the brain to help suppress arousability. Higher protein concentration in the gut leads to increased activity of enteroendocrine cells that release the peptide CCHa1. CCHa1 signals to a small group of dopamine neurons in the brain to modulate their activity; the dopaminergic activity regulates the behavioral responsiveness of animals to vibrations. The CCHa1 pathway and dietary proteins do not influence responsiveness to all sensory inputs, showing that during sleep, different information streams can be gated through independent mechanisms.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no conflict of interest.
Figures
Comment in
-
Light sleeper? Eat more protein.Sci Signal. 2023 Apr 4;16(779):eadh9952. doi: 10.1126/scisignal.adh9952. Epub 2023 Apr 4. Sci Signal. 2023. PMID: 37014926
-
Do flies dream of protein? How the gut regulates sleep depth.Cell Metab. 2023 May 2;35(5):732-734. doi: 10.1016/j.cmet.2023.04.001. Cell Metab. 2023. PMID: 37137287
-
Sleep regulation: The gut sets the threshold.Curr Biol. 2023 May 22;33(10):R420-R423. doi: 10.1016/j.cub.2023.04.014. Curr Biol. 2023. PMID: 37220738
References
-
- Kline C. (2013). Sleep Quality. In Encyclopedia of Behavioral Medicine (Springer New York; ), pp. 1811–1813. 10.1007/978-1-4419-1005-9_849. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
