

A transcriptionally distinct subset of influenza-specific effector memory B cells predicts long-lived antibody responses to vaccination in humans
- PMID: 36958335
- PMCID: PMC10113805
- DOI: 10.1016/j.immuni.2023.03.001
A transcriptionally distinct subset of influenza-specific effector memory B cells predicts long-lived antibody responses to vaccination in humans
Abstract
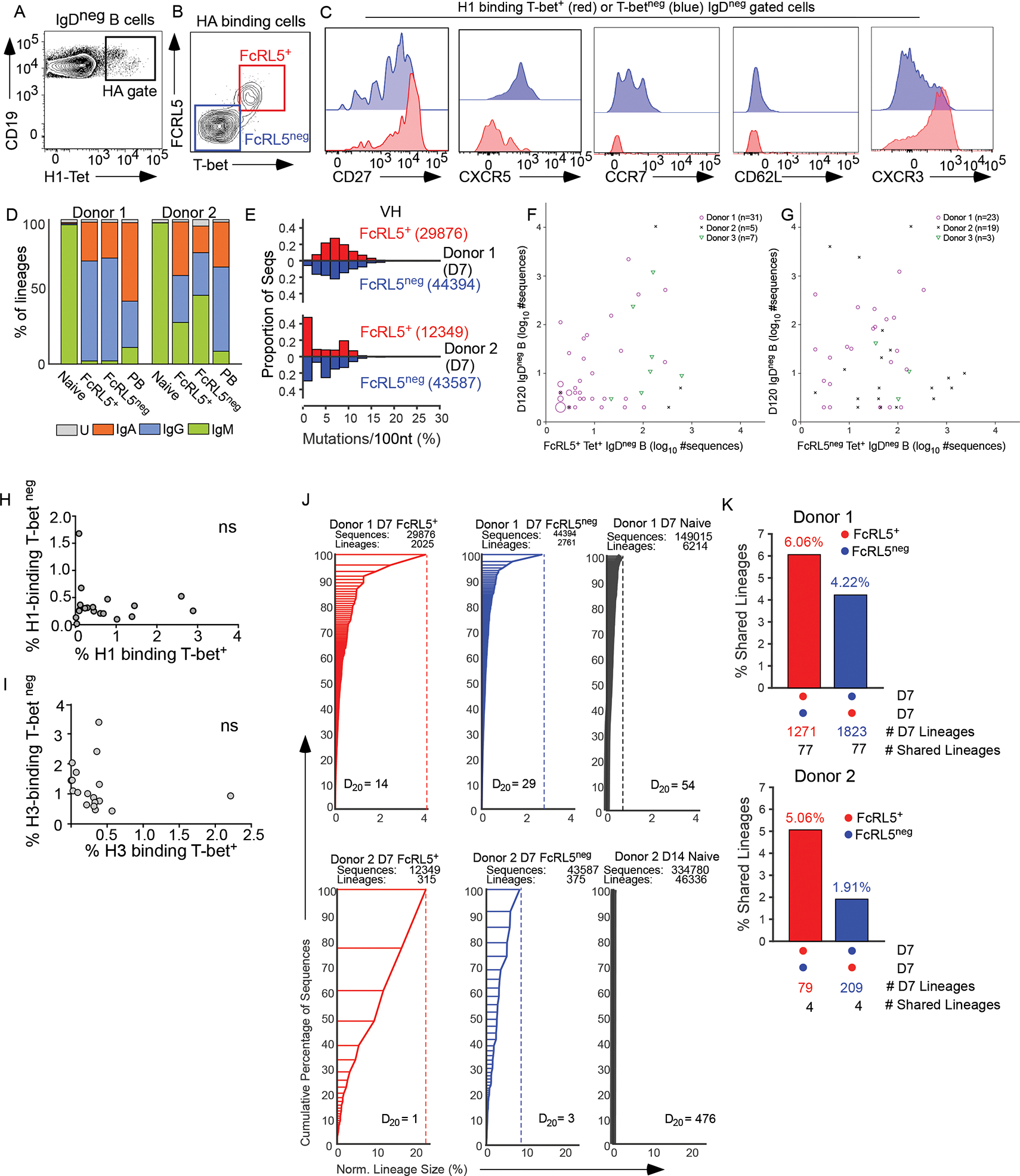
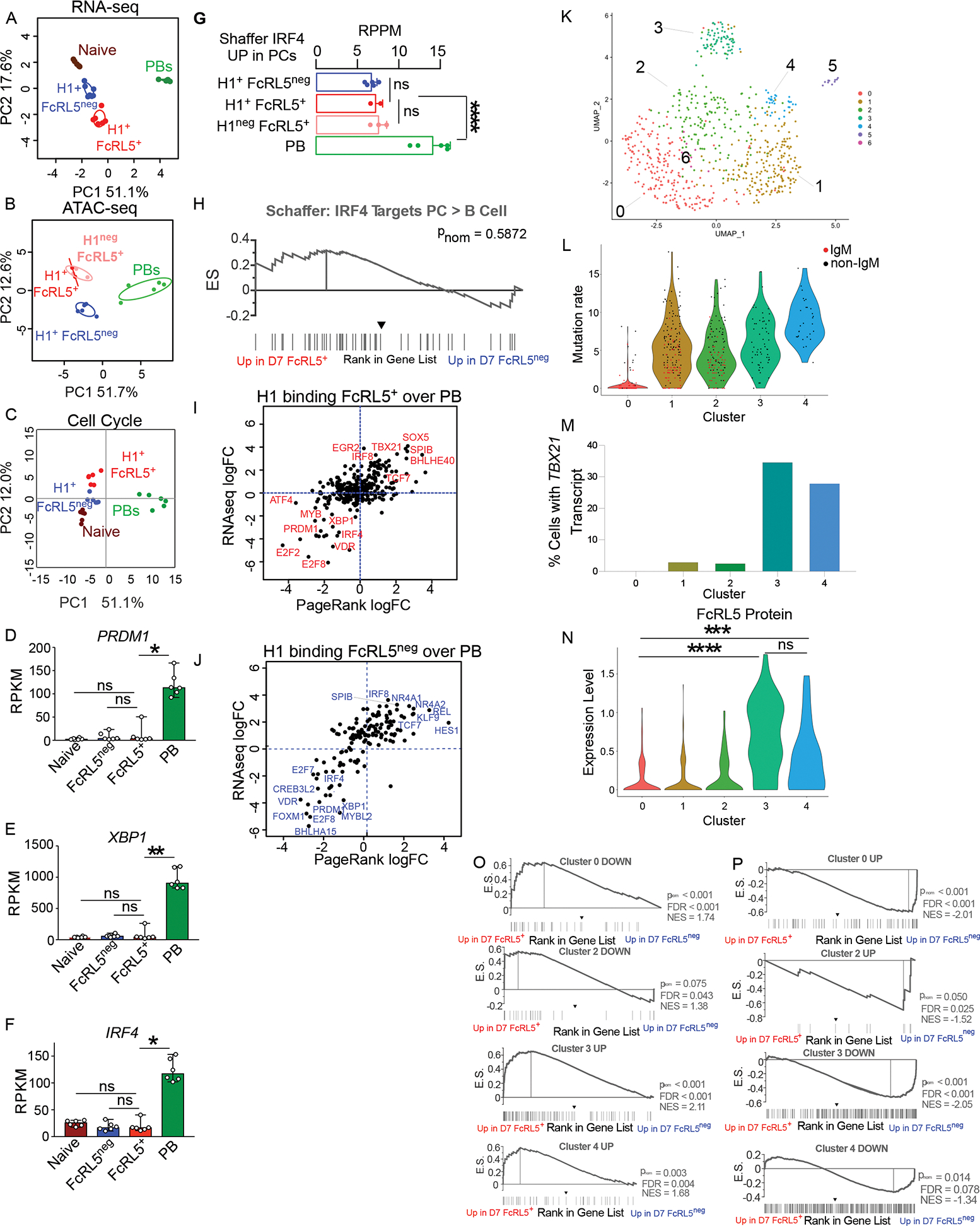
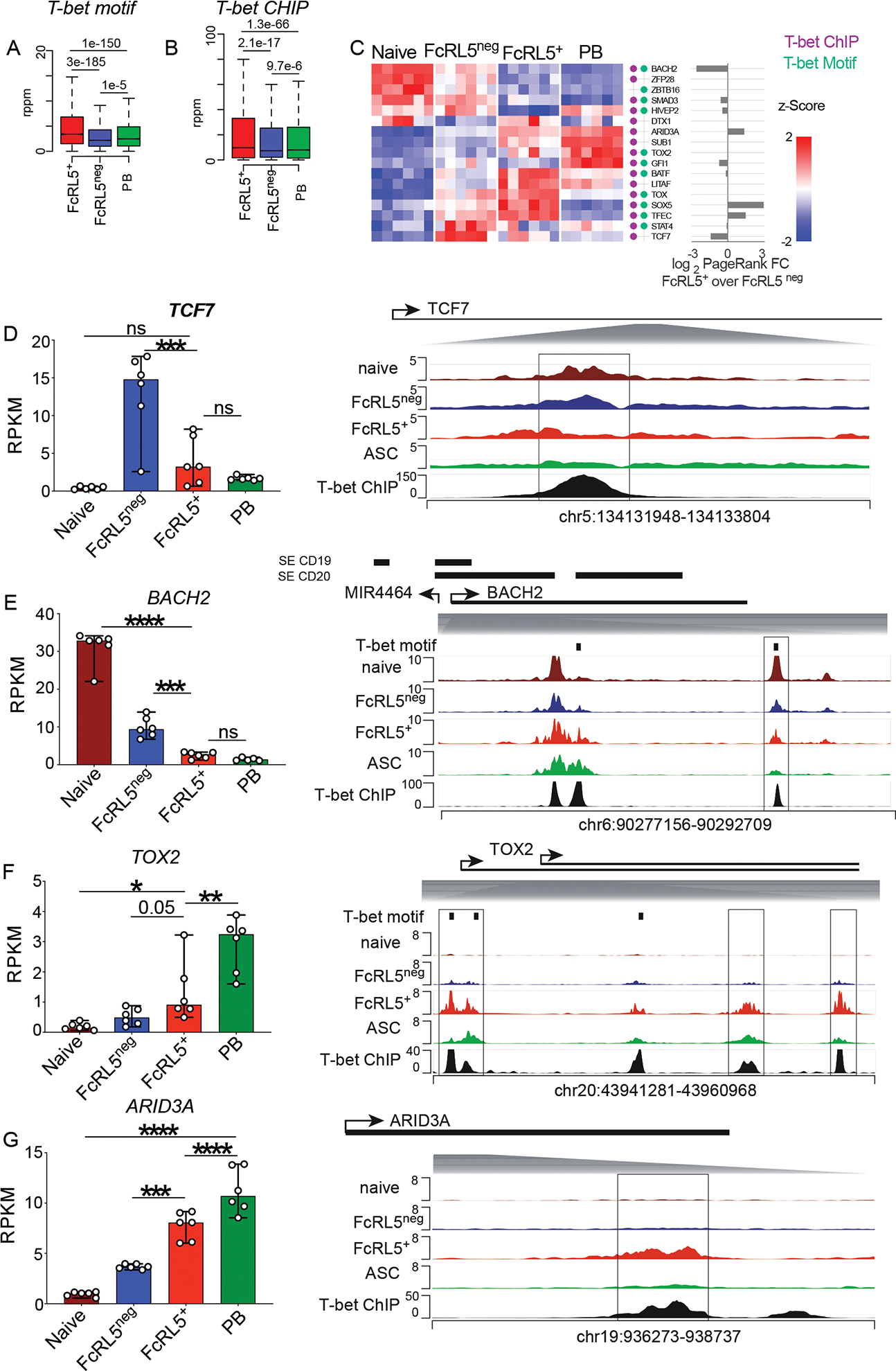
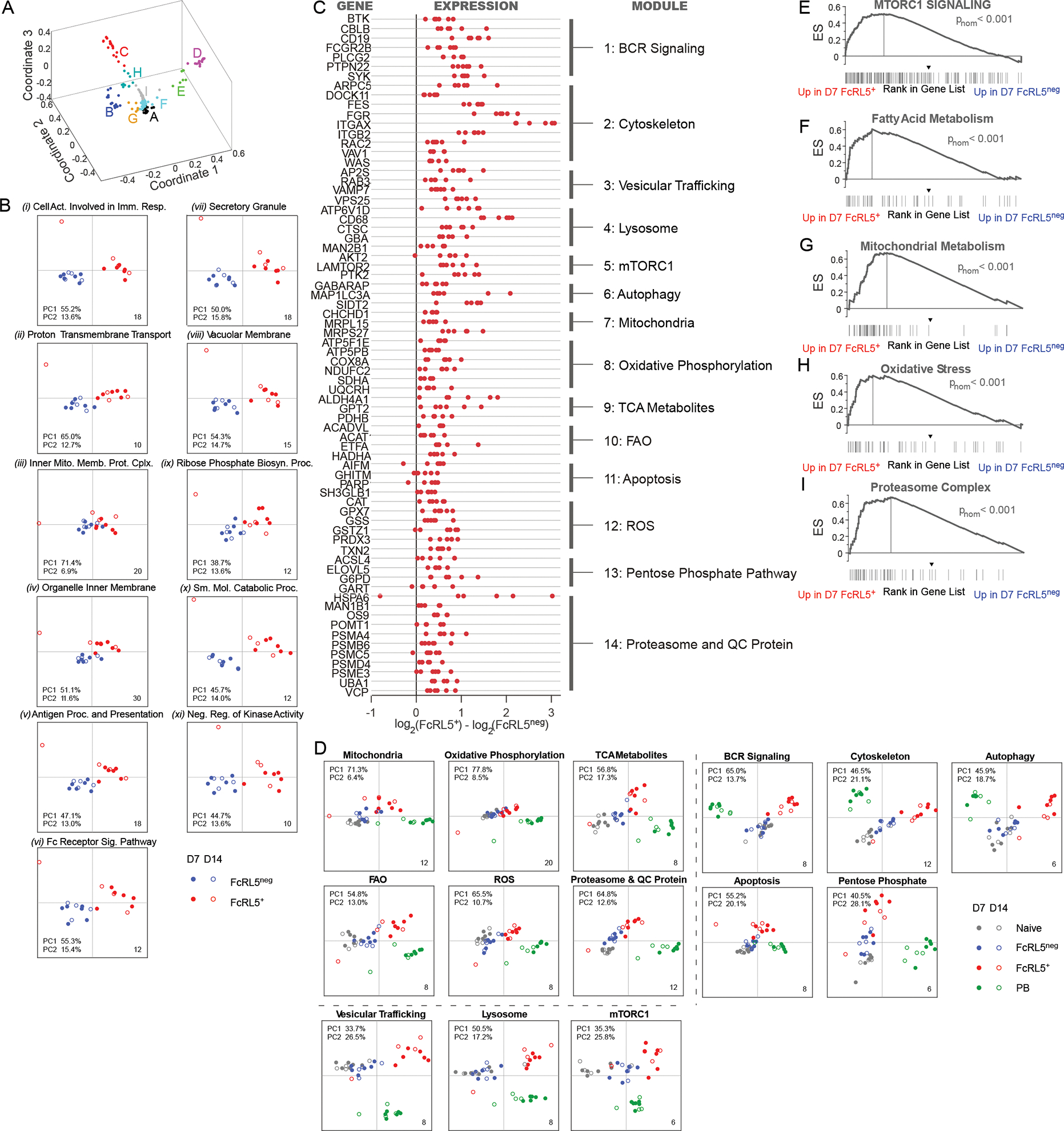
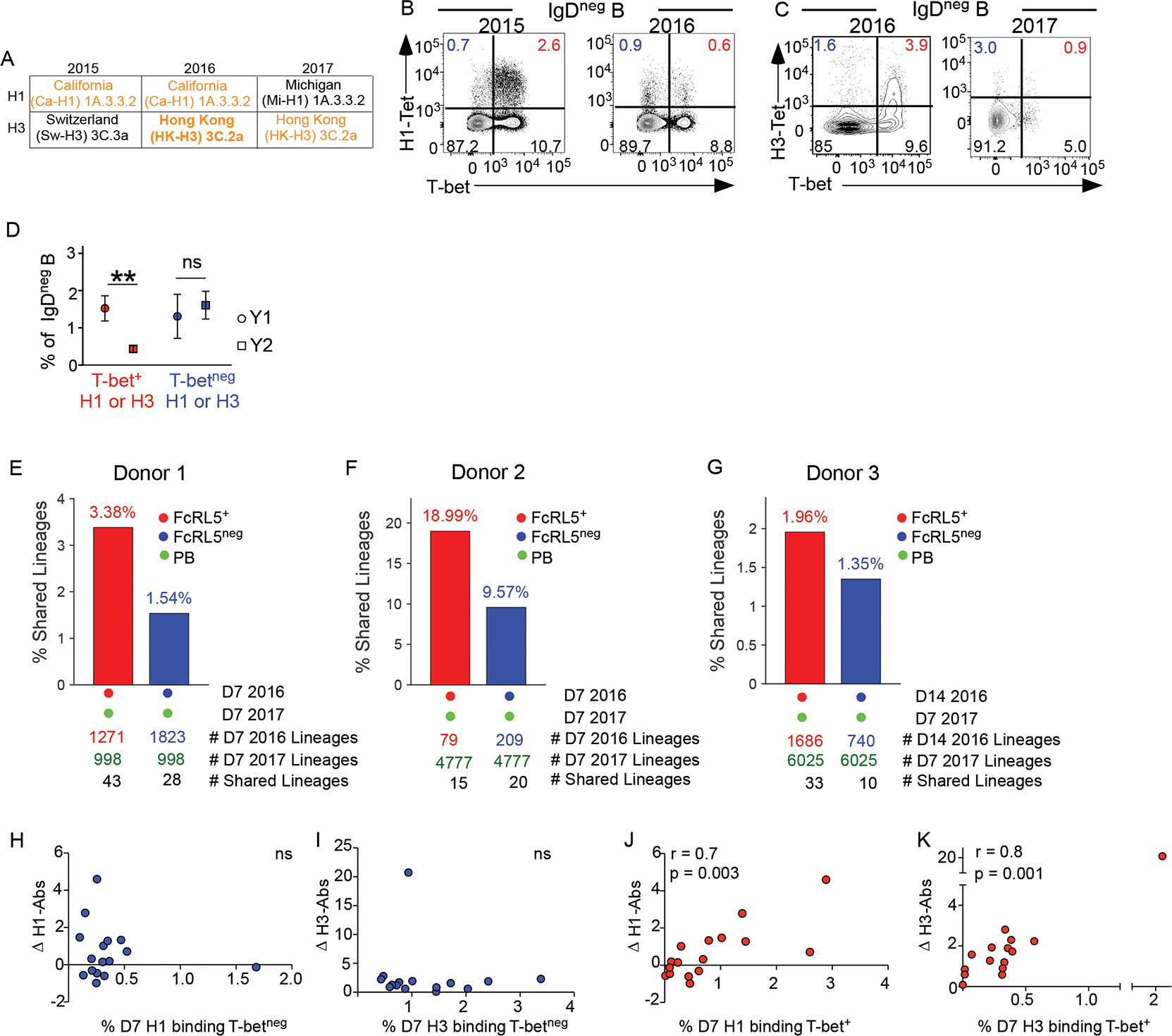
Seasonal influenza vaccination elicits hemagglutinin (HA)-specific memory B (Bmem) cells, and although multiple Bmem cell populations have been characterized, considerable heterogeneity exists. We found that HA-specific human Bmem cells differed in the expression of surface marker FcRL5 and transcriptional factor T-bet. FcRL5+T-bet+ Bmem cells were transcriptionally similar to effector-like memory cells, while T-betnegFcRL5neg Bmem cells exhibited stem-like central memory properties. FcRL5+ Bmem cells did not express plasma-cell-commitment factors but did express transcriptional, epigenetic, metabolic, and functional programs that poised these cells for antibody production. Accordingly, HA+ T-bet+ Bmem cells at day 7 post-vaccination expressed intracellular immunoglobulin, and tonsil-derived FcRL5+ Bmem cells differentiated more rapidly into antibody-secreting cells (ASCs) in vitro. The T-bet+ Bmem cell response positively correlated with long-lived humoral immunity, and clonotypes from T-bet+ Bmem cells were represented in the secondary ASC response to repeat vaccination, suggesting that this effector-like population predicts influenza vaccine durability and recall potential.
Keywords: T-bet; durable immunity; influenza vaccination; memory B cells; plasmablasts.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures


References
-
- Lee FE, Halliley JL, Walsh EE, Moscatiello AP, Kmush BL, Falsey AR, Randall TD, Kaminiski DA, Miller RK, and Sanz I (2011). Circulating human antibody-secreting cells during vaccinations and respiratory viral infections are characterized by high specificity and lack of bystander effect. J Immunol 186, 5514–5521. 10.4049/jimmunol.1002932. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
