

Co-option of epidermal cells enables touch sensing
- PMID: 36959505
- PMCID: PMC10104782
- DOI: 10.1038/s41556-023-01110-2
Co-option of epidermal cells enables touch sensing
Erratum in
-
Author Correction: Co-option of epidermal cells enables touch sensing.Nat Cell Biol. 2024 Jan;26(1):170. doi: 10.1038/s41556-023-01326-2. Nat Cell Biol. 2024. PMID: 38057364 Free PMC article. No abstract available.
Abstract
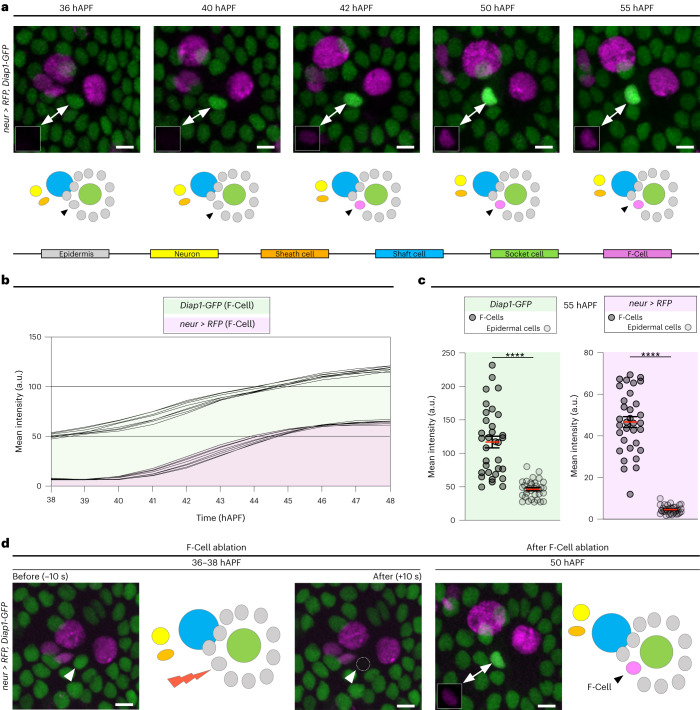
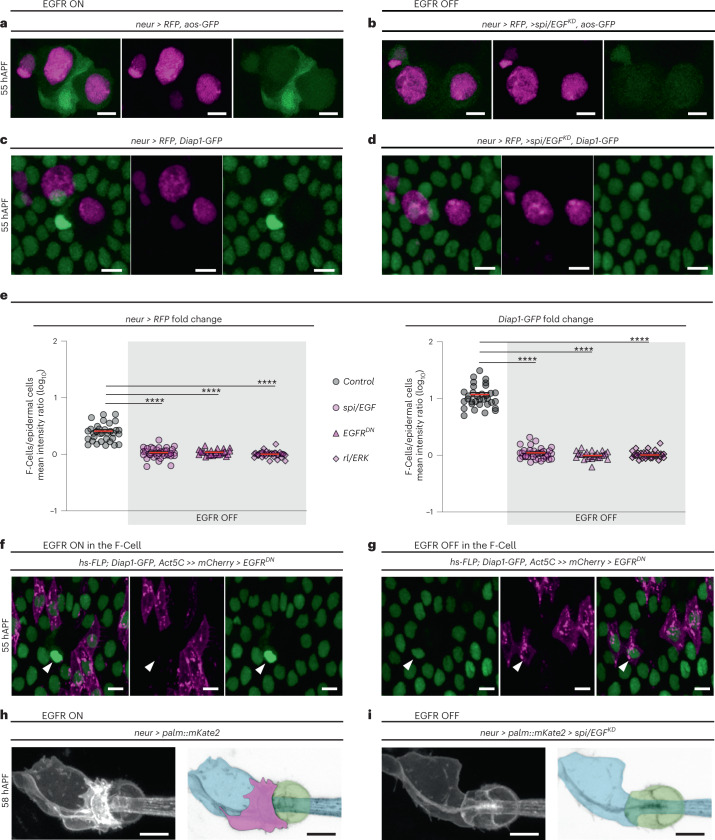
The epidermis is equipped with specialized mechanosensory organs that enable the detection of tactile stimuli. Here, by examining the differentiation of the tactile bristles, mechanosensory organs decorating the Drosophila adult epidermis, we show that neighbouring epidermal cells are essential for touch perception. Each mechanosensory bristle signals to the surrounding epidermis to co-opt a single epidermal cell, which we named the F-Cell. Once specified, the F-Cell adopts a specialized morphology to ensheath each bristle. Functional assays reveal that adult mechanosensory bristles require association with the epidermal F-Cell for touch sensing. Our findings underscore the importance of resident epidermal cells in the assembly of functional touch-sensitive organs.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures














Comment in
-
A new cell in an old tactile sensory organ.Nat Cell Biol. 2023 Apr;25(4):518-519. doi: 10.1038/s41556-023-01119-7. Nat Cell Biol. 2023. PMID: 37059881 No abstract available.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
