

Effect of peripheral cellular senescence on brain aging and cognitive decline
- PMID: 36959691
- PMCID: PMC10186609
- DOI: 10.1111/acel.13817
Effect of peripheral cellular senescence on brain aging and cognitive decline
Abstract
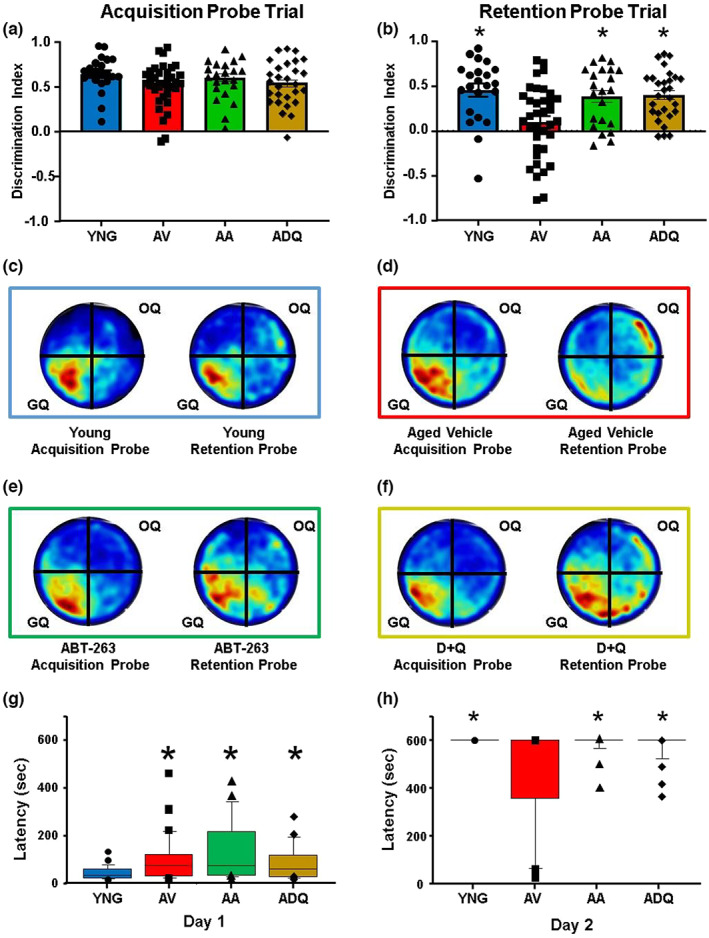
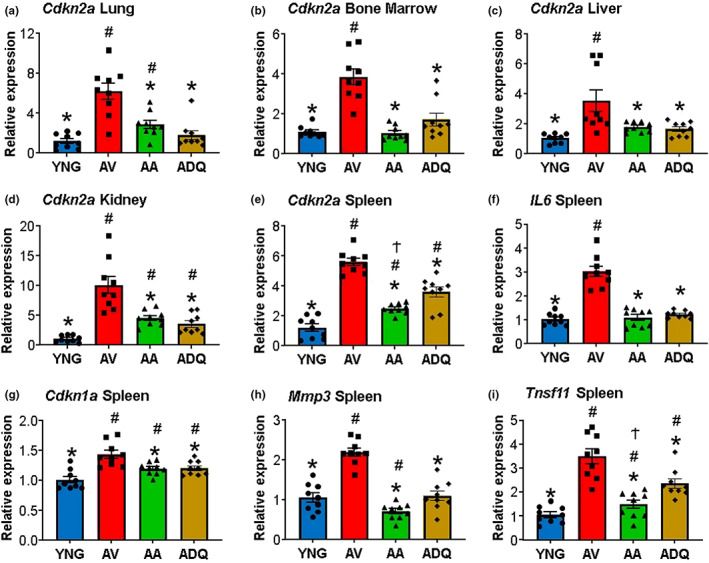
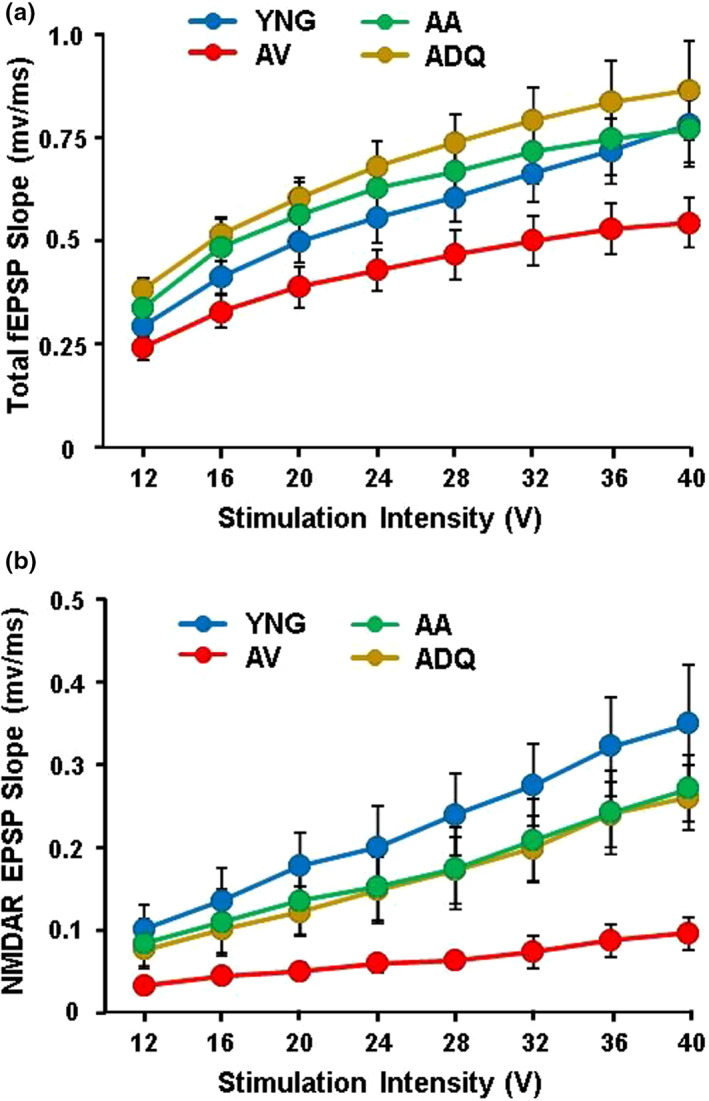
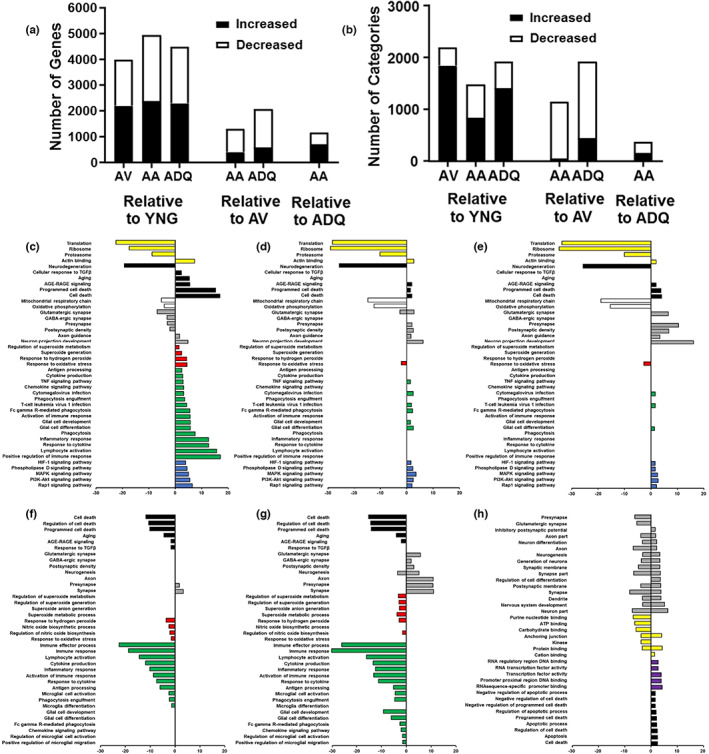
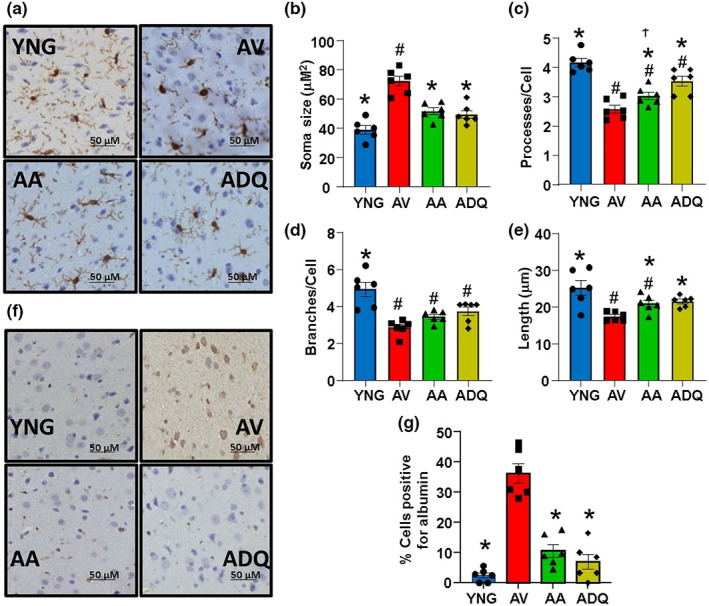
We examine similar and differential effects of two senolytic treatments, ABT-263 and dasatinib + quercetin (D + Q), in preserving cognition, markers of peripheral senescence, and markers of brain aging thought to underlie cognitive decline. Male F344 rats were treated from 12 to 18 months of age with D + Q, ABT-263, or vehicle, and were compared to young (6 months). Both senolytic treatments rescued memory, preserved the blood-brain barrier (BBB) integrity, and prevented the age-related decline in hippocampal N-methyl-D-aspartate receptor (NMDAR) function associated with impaired cognition. Senolytic treatments decreased senescence-associated secretory phenotype (SASP) and inflammatory cytokines/chemokines in the plasma (IL-1β, IP-10, and RANTES), with some markers more responsive to D + Q (TNFα) or ABT-263 (IFNγ, leptin, EGF). ABT-263 was more effective in decreasing senescence genes in the spleen. Both senolytic treatments decreased the expression of immune response and oxidative stress genes and increased the expression of synaptic genes in the dentate gyrus (DG). However, D + Q influenced twice as many genes as ABT-263. Relative to D + Q, the ABT-263 group exhibited increased expression of DG genes linked to cell death and negative regulation of apoptosis and microglial cell activation. Furthermore, D + Q was more effective at decreasing morphological markers of microglial activation. The results indicate that preserved cognition was associated with the removal of peripheral senescent cells, decreasing systemic inflammation that normally drives neuroinflammation, BBB breakdown, and impaired synaptic function. Dissimilarities associated with brain transcription indicate divergence in central mechanisms, possibly due to differential access.
Keywords: aging; cognition; inflammation; oxidative stress; senolytic NMDA receptor; transcription.
© 2023 The Authors. Aging Cell published by Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
Daohong Zhou is an inventor of two pending patent applications for use of Bcl‐xL PROTACs, synthesized using ABT‐263, as senolytic and antitumor agents. Daohong Zhou is a co‐founder of and has equity in Dialectic Therapeutics, which develops Bcl‐xL PROTACs to treat cancer.
Figures

References
-
- Balusu, S. , van Wonterghem, E. , de Rycke, R. , Raemdonck, K. , Stremersch, S. , Gevaert, K. , Brkic, M. , Demeestere, D. , Vanhooren, V. , Hendrix, A. , Libert, C. , & Vandenbroucke, R. E. (2016). Identification of a novel mechanism of blood‐brain communication during peripheral inflammation via choroid plexus‐derived extracellular vesicles. EMBO Molecular Medicine, 8(10), 1162–1183. 10.15252/emmm.201606271 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
