

A novel feature of the ancient organ: A possible involvement of the subcommissural organ in neurogenic/gliogenic potential in the adult brain
- PMID: 36960167
- PMCID: PMC10027738
- DOI: 10.3389/fnins.2023.1141913
A novel feature of the ancient organ: A possible involvement of the subcommissural organ in neurogenic/gliogenic potential in the adult brain
Abstract
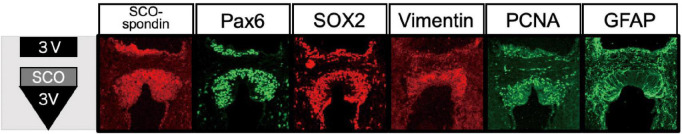
The subcommissural organ (SCO) is a circumventricular organ highly conserved in vertebrates from Cyclostomata such as lamprey to mammals including human. The SCO locates in the boundary between the third ventricle and the entrance of the aqueduct of Sylvius. The SCO functions as a secretory organ producing a variety of proteins such as SCO-spondin, transthyretin, and basic fibroblast growth factor (FGF) into the cerebrospinal fluid (CSF). A significant contribution of the SCO has been thought to maintain the homeostasis of CSF dynamics. However, evidence has shown a possible role of SCO on neurogenesis in the adult brain. This review highlights specific features of the SCO related to adult neurogenesis, suggested by the progress of understanding SCO functions. We begin with a brief history of the SCO discovery and continue to structural features, gene expression, and a possible role in adult neurogenesis suggested by the SCO transplant experiment.
Keywords: Pax6; Sox2; neural stem progenitor cell; neurogenesis; subcommissural organ.
Copyright © 2023 Inada, Corales and Osumi.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures



References
-
- Blackshear P. J., Graves J. P., Stumpo D. J., Cobos I., Rubenstein J. L., Zeldin D. C. (2003). Graded phenotypic response to partial and complete deficiency of a brain-specific transcript variant of the winged helix transcription factor RFX4. Development 130 4539–4552. 10.1242/dev.00661 - DOI - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Research Materials