

CsTRM5 regulates fruit shape via mediating cell division direction and cell expansion in cucumber
- PMID: 36960430
- PMCID: PMC10028494
- DOI: 10.1093/hr/uhad007
CsTRM5 regulates fruit shape via mediating cell division direction and cell expansion in cucumber
Abstract
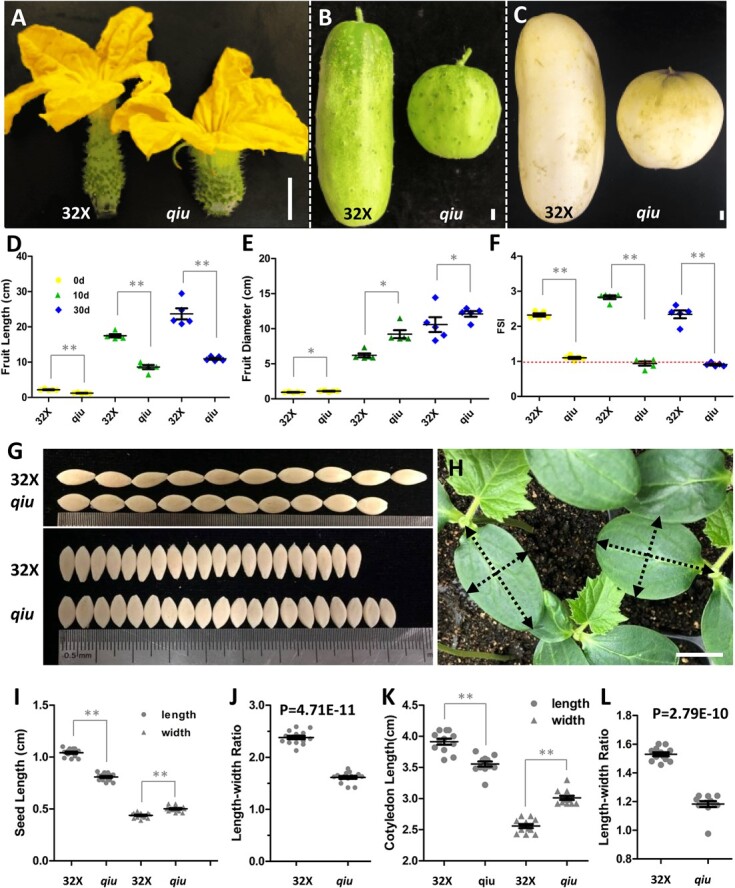
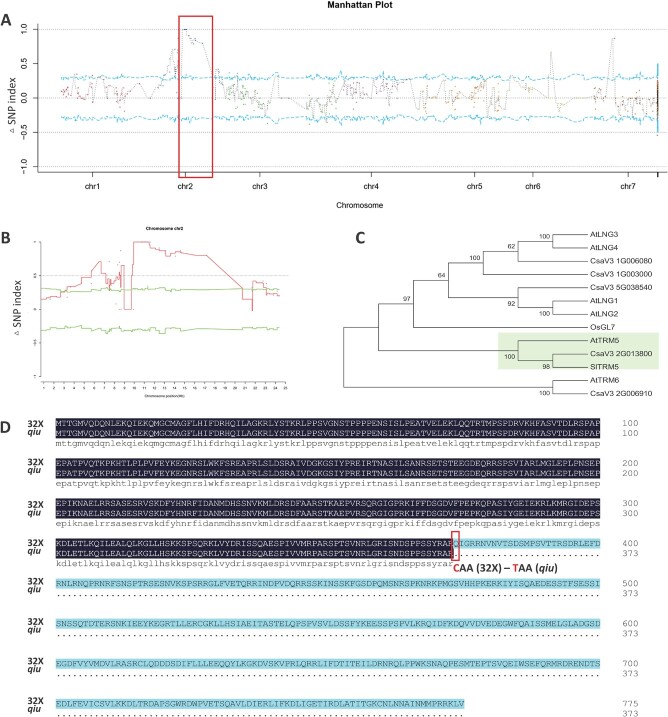
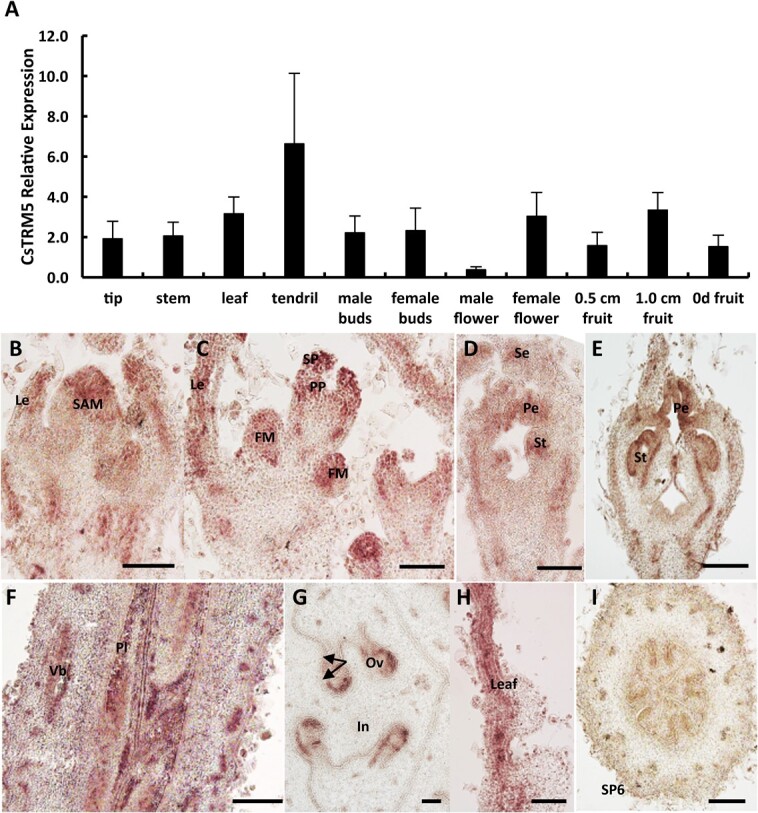
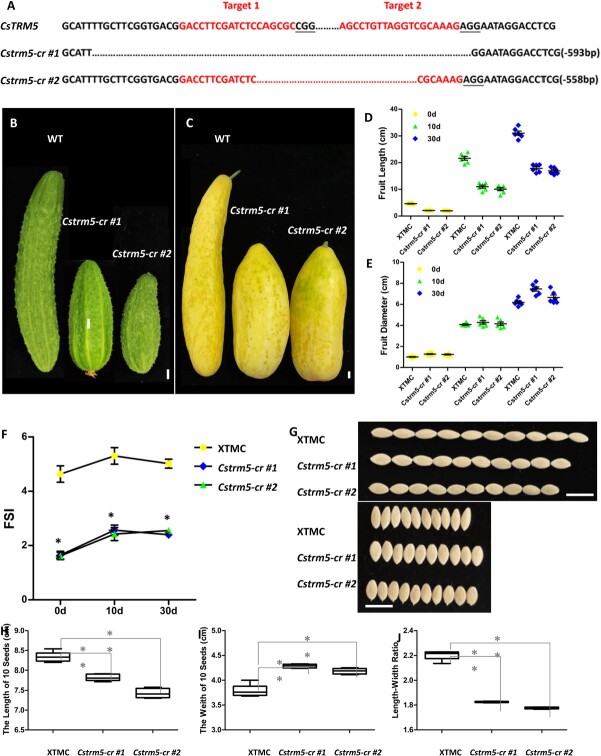
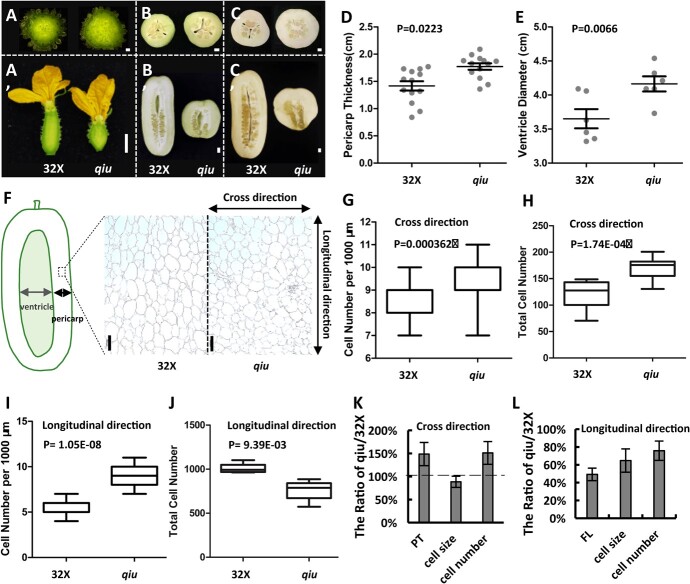
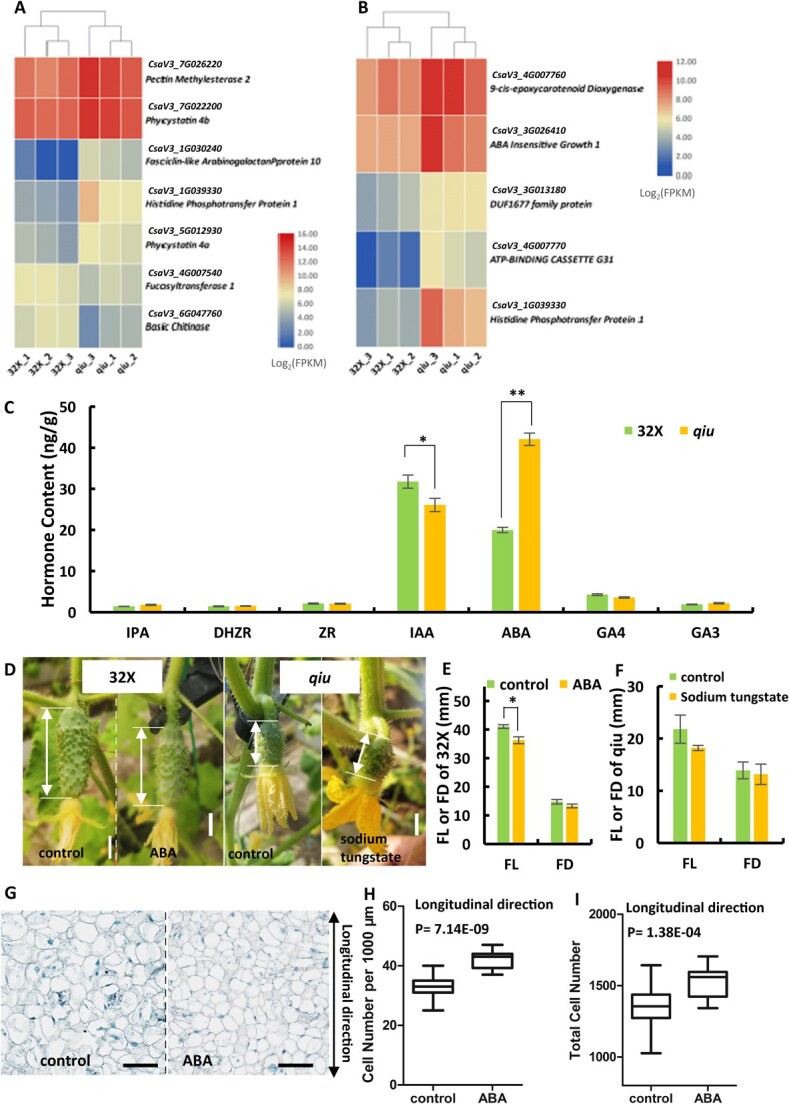
Fruit shape and size are important appearance and yield traits in cucumber, but the underlying genes and their regulatory mechanisms remain poorly understood. Here we identified a mutant with spherical fruits from an Ethyl Methane Sulfonate (EMS)-mutagenized library, named the qiu mutant. Compared with the cylindrical fruit shape in 32X (wild type), the fruit shape in qiu was round due to reduced fruit length and increased fruit diameter. MutMap analysis narrowed the candidate gene in the 6.47 MB range on Chr2, harboring the FS2.1 locus reported previously. A single-nucleotide polymorphism (SNP) (11359603) causing a truncated protein of CsaV3_2G013800, the homolog of tomato fruit shape gene SlTRM5, may underlie the fruit shape variation in the qiu mutant. Knockout of CsTRM5 by the CRISPR-Cas9 system confirmed that CsaV3_2G013800/CsTRM5 was the causal gene responsible for qiu. Sectioning analysis showed that the spherical fruit in qiu resulted mainly from increased and reduced cell division along the transverse and longitudinal directions, respectively. Meanwhile, the repressed cell expansion contributed to the decreased fruit length in qiu. Transcriptome profiling showed that the expression levels of cell-wall-related genes and abscisic acid (ABA) pathway genes were significantly upregulated in qiu. Hormone measurements indicated that ABA content was greatly increased in the qiu mutant. Exogenous ABA application reduced fruit elongation by inhibiting cell expansion in cucumber. Taken together, these data suggest that CsTRM5 regulates fruit shape by affecting cell division direction and cell expansion, and that ABA participates in the CsTRM5-mediated cell expansion during fruit elongation in cucumber.
© The Author(s) 2023. Published by Oxford University Press on behalf of Nanjing Agricultural University.
Conflict of interest statement
The authors declare no conflict of interests.
Figures
References
-
- Li S, Pan Y, Wen Cet al. Integrated analysis in bi-parental and natural populations reveals CsCLAVATA3 (CsCLV3) underlying carpel number variations in cucumber. Theor Appl Genet. 2016;129:1007–22. - PubMed
-
- Pan Y, Wang Y, McGregor Cet al. Genetic architecture of fruit size and shape variation in cucurbits: a comparative perspective. Theor Appl Genet. 2020;133:1–21. - PubMed
-
- Che G, Zhang X. Molecular basis of cucumber fruit domestication. Curr Opin Plant Biol. 2019;47:38–46. - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
