

Enterobacterales plasmid sharing amongst human bloodstream infections, livestock, wastewater, and waterway niches in Oxfordshire, UK
- PMID: 36961866
- PMCID: PMC10703447
- DOI: 10.7554/eLife.85302
Enterobacterales plasmid sharing amongst human bloodstream infections, livestock, wastewater, and waterway niches in Oxfordshire, UK
Abstract
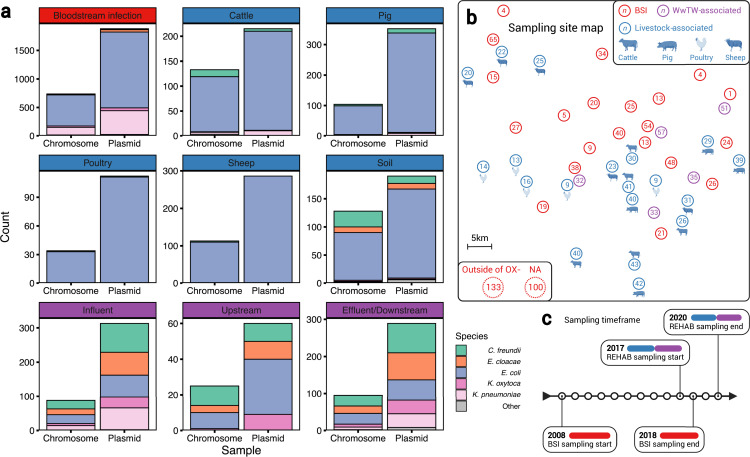
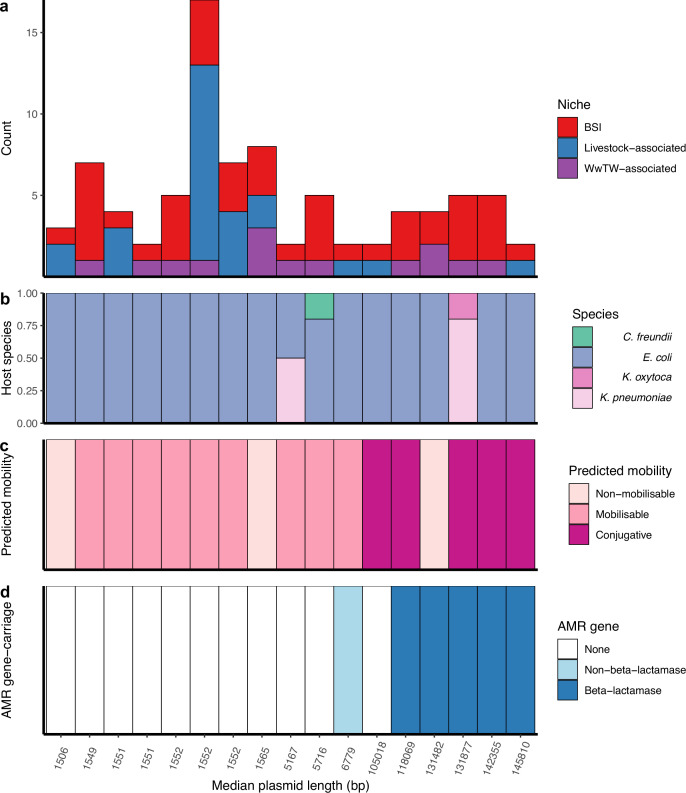
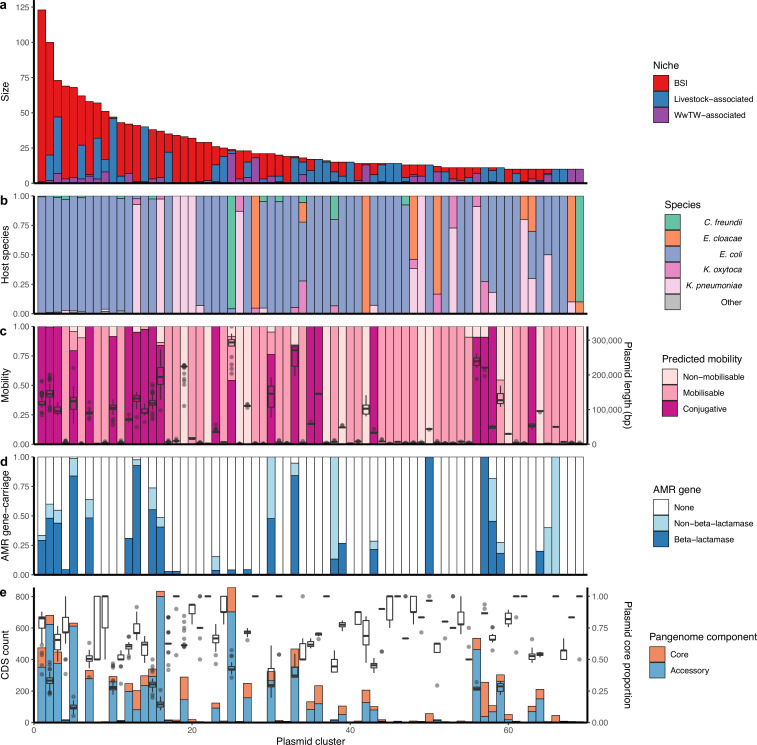
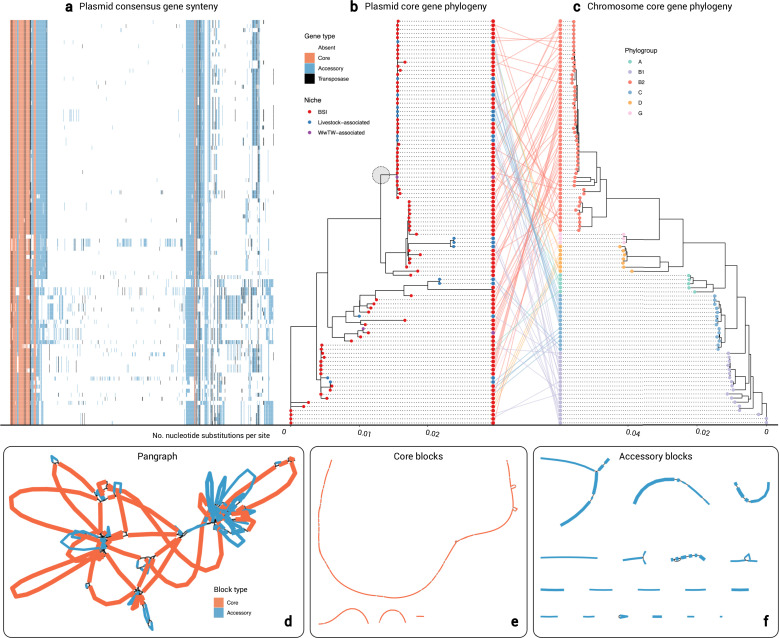
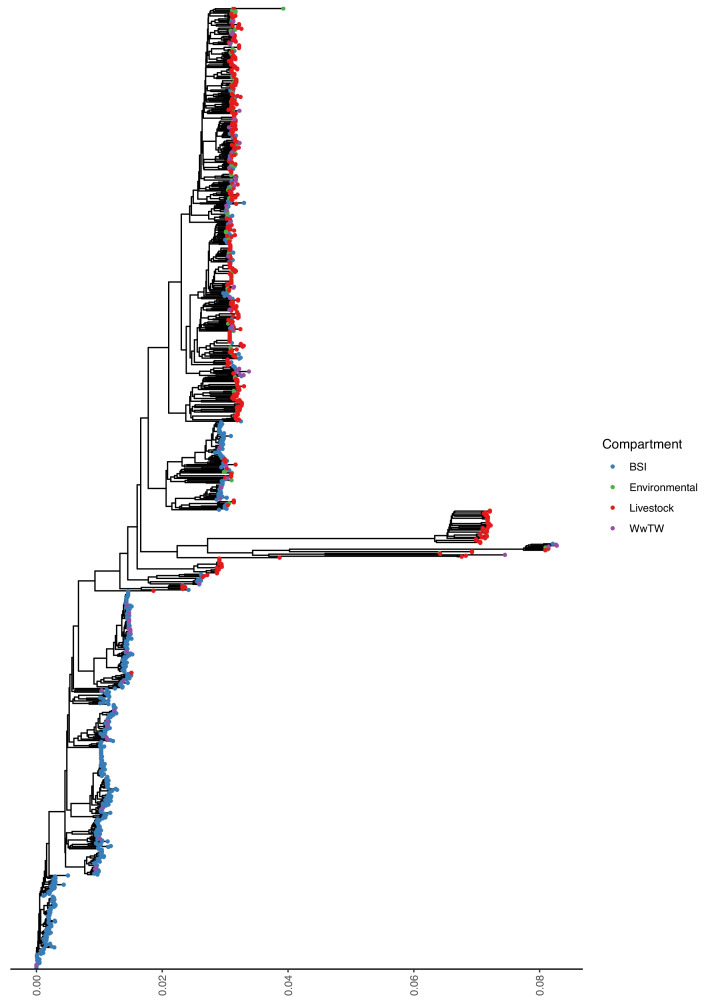
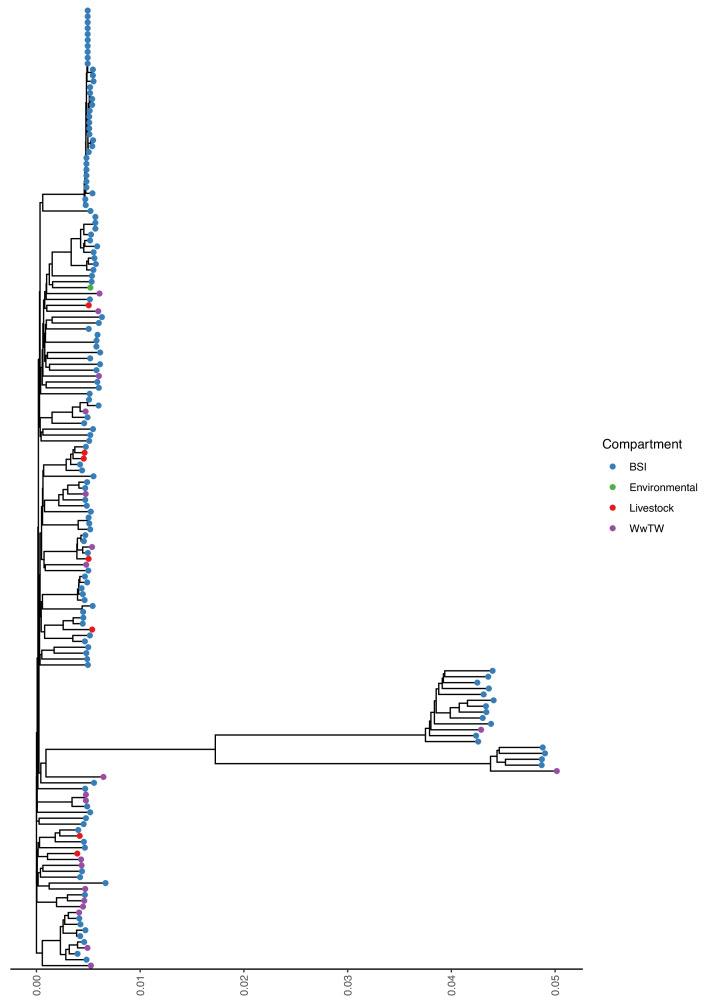
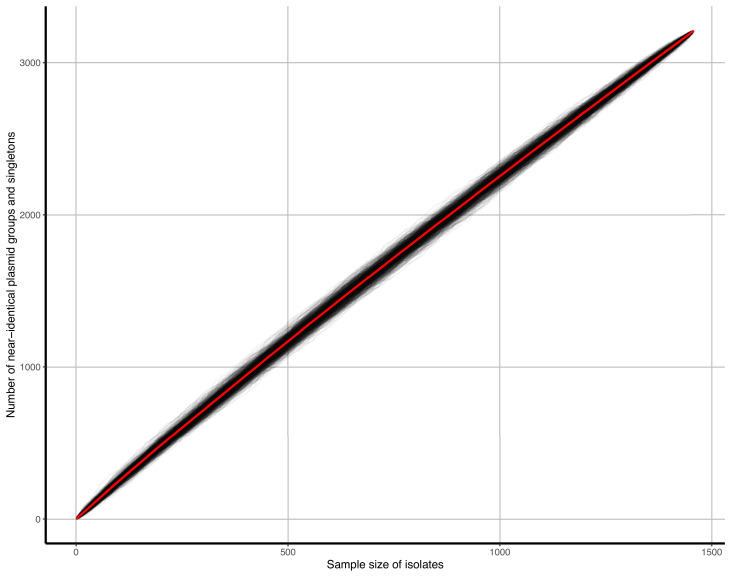
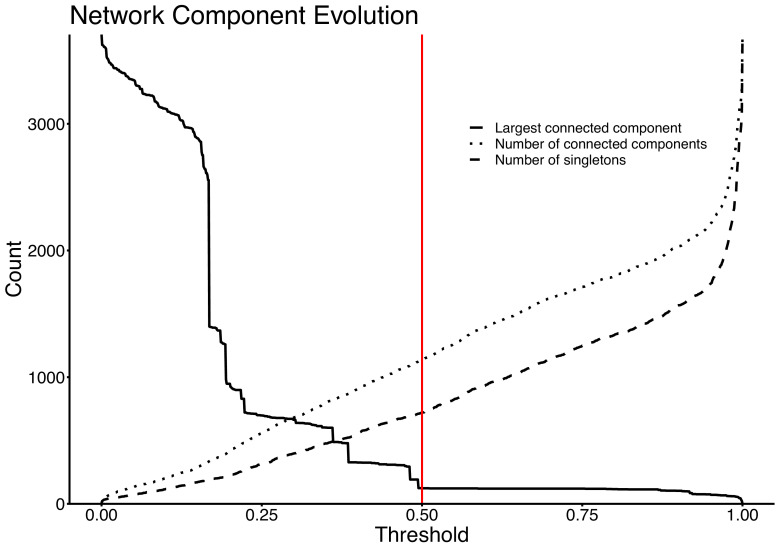
Plasmids enable the dissemination of antimicrobial resistance (AMR) in common Enterobacterales pathogens, representing a major public health challenge. However, the extent of plasmid sharing and evolution between Enterobacterales causing human infections and other niches remains unclear, including the emergence of resistance plasmids. Dense, unselected sampling is essential to developing our understanding of plasmid epidemiology and designing appropriate interventions to limit the emergence and dissemination of plasmid-associated AMR. We established a geographically and temporally restricted collection of human bloodstream infection (BSI)-associated, livestock-associated (cattle, pig, poultry, and sheep faeces, farm soils) and wastewater treatment work (WwTW)-associated (influent, effluent, waterways upstream/downstream of effluent outlets) Enterobacterales. Isolates were collected between 2008 and 2020 from sites <60 km apart in Oxfordshire, UK. Pangenome analysis of plasmid clusters revealed shared 'backbones', with phylogenies suggesting an intertwined ecology where well-conserved plasmid backbones carry diverse accessory functions, including AMR genes. Many plasmid 'backbones' were seen across species and niches, raising the possibility that plasmid movement between these followed by rapid accessory gene change could be relatively common. Overall, the signature of identical plasmid sharing is likely to be a highly transient one, implying that plasmid movement might be occurring at greater rates than previously estimated, raising a challenge for future genomic One Health studies.
Keywords: E. coli; Enterobacterales; K. pneumoniae; One Health; antimicrobial resistance; bloodstream infections; genetics; genomic epidemiology; genomics; infectious disease; microbiology; plasmid.
© 2023, Matlock, Lipworth et al.
Conflict of interest statement
WM, SL, KC, MA, LB, JK, MA, SO, MM, DC, DR, MA, LS, NS No competing interests declared
Figures
Update of
References
-
- AbuOun M, Jones H, Stubberfield E, Gilson D, Shaw LP, Hubbard ATM, Chau KK, Sebra R, Peto TEA, Crook DW, Read DS, Gweon HS, Walker AS, Stoesser N, Smith RP, Anjum MF, On Behalf Of The Rehab Consortium A genomic epidemiological study shows that prevalence of antimicrobial resistance in enterobacterales is associated with the livestock host, as well as antimicrobial usage. Microbial Genomics. 2021;7:000630. doi: 10.1099/mgen.0.000630. - DOI - PMC - PubMed
-
- Bastidas-Caldes C, de Waard JH, Salgado MS, Villacís MJ, Coral-Almeida M, Yamamoto Y, Calvopiña M. Worldwide prevalence of mcr-mediated colistin-resistance Escherichia coli in isolates of clinical samples, healthy humans, and livestock-A systematic review and meta-analysis. Pathogens. 2022;11:659. doi: 10.3390/pathogens11060659. - DOI - PMC - PubMed
-
- Blondel VD, Guillaume JL, Lambiotte R, Lefebvre E. Fast unfolding of communities in large networks. Journal of Statistical Mechanics. 2008;2008:10008. doi: 10.1088/1742-5468/2008/10/P10008. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
