

The transcription factor Mef2d regulates B:T synapse-dependent GC-TFH differentiation and IL-21-mediated humoral immunity
- PMID: 36961907
- PMCID: PMC10311795
- DOI: 10.1126/sciimmunol.adf2248
The transcription factor Mef2d regulates B:T synapse-dependent GC-TFH differentiation and IL-21-mediated humoral immunity
Abstract
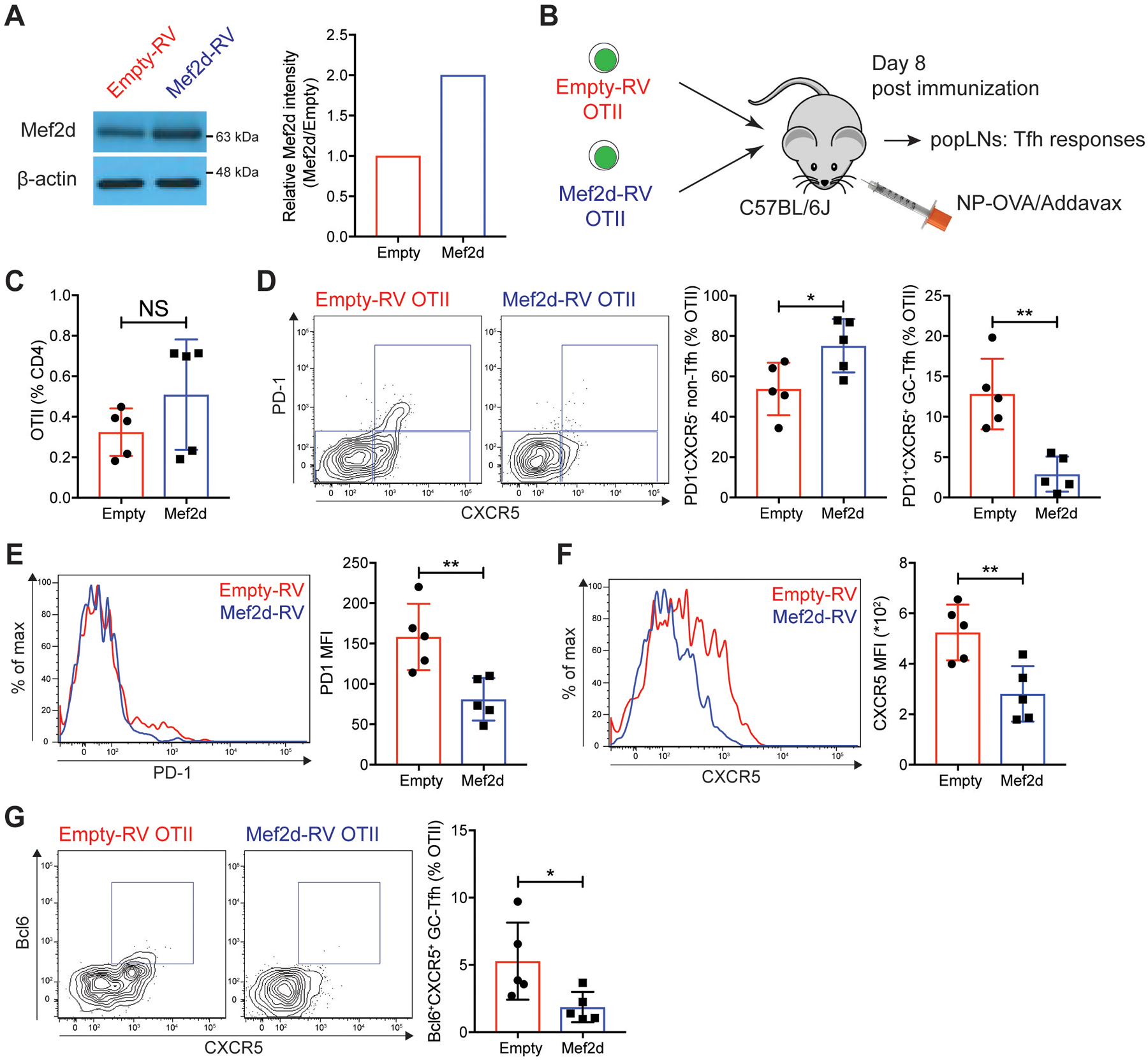
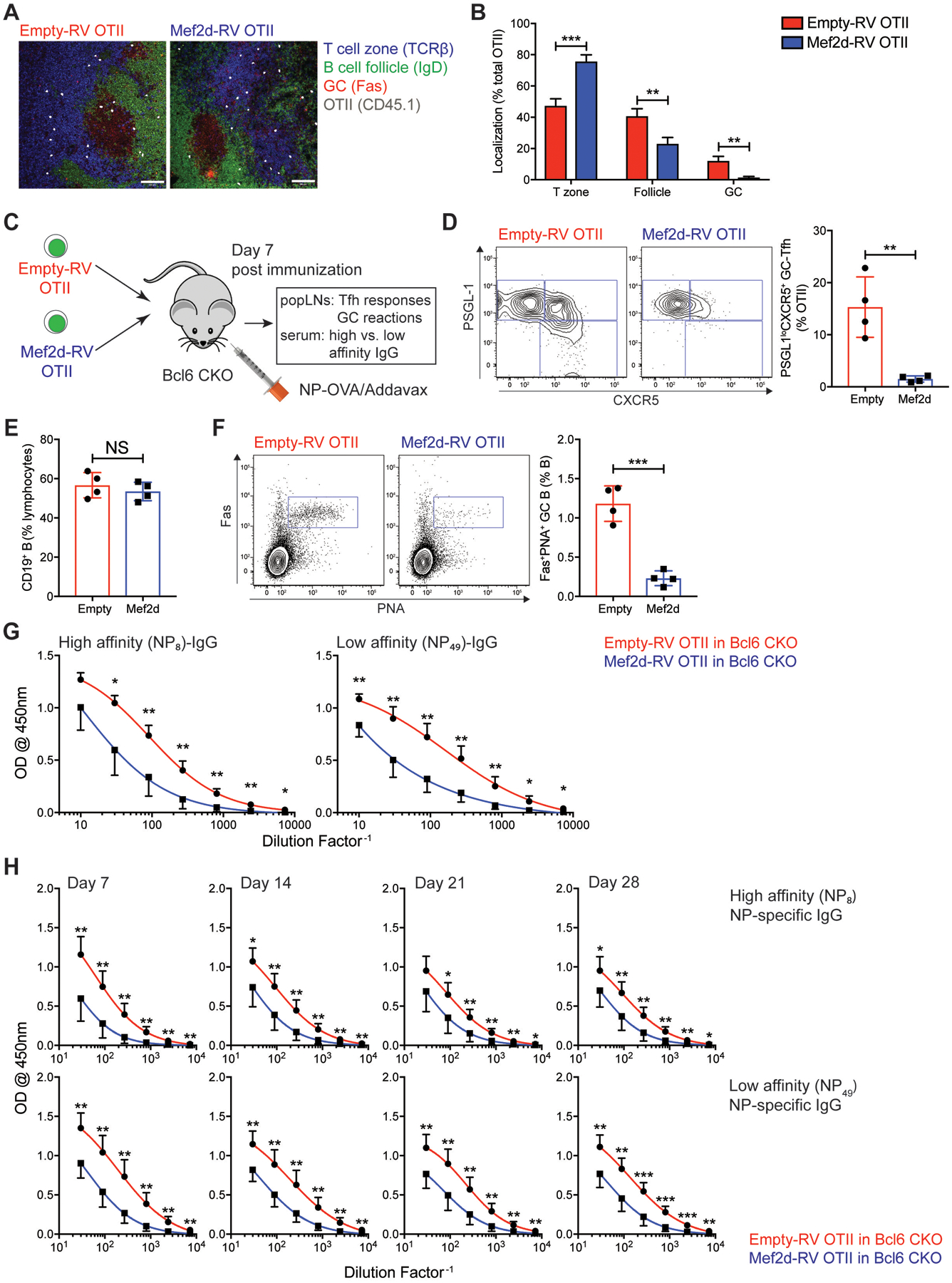
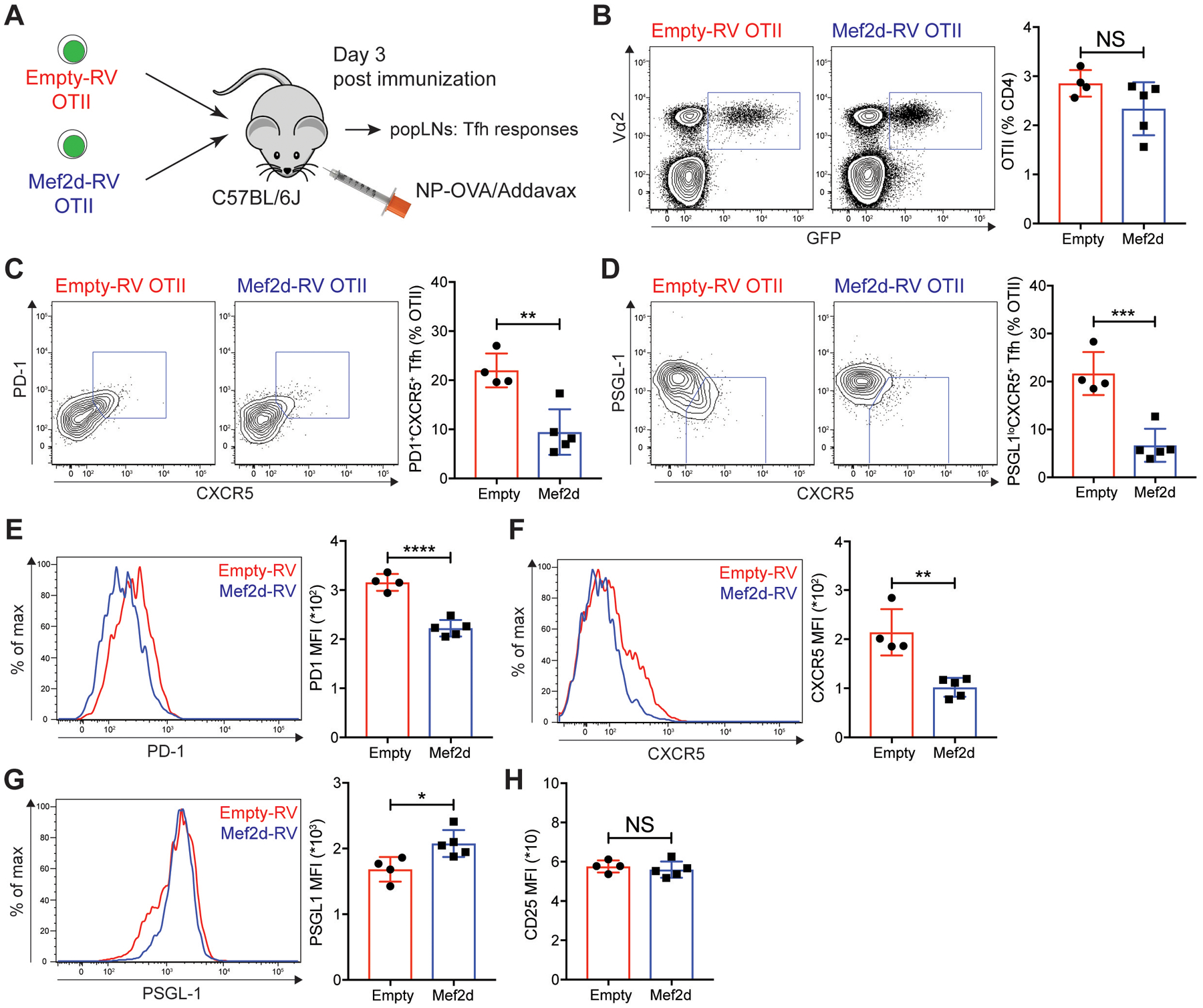
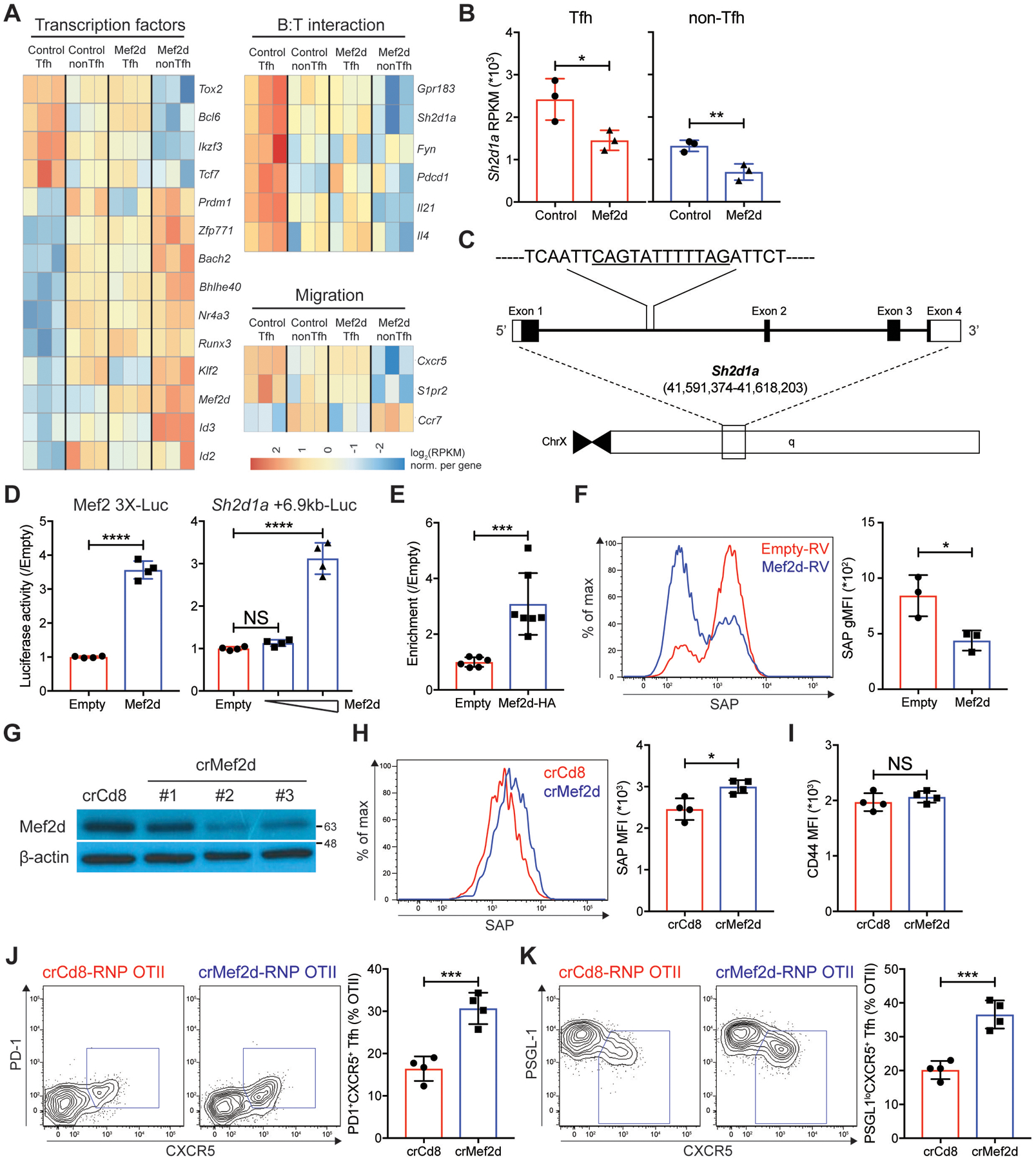
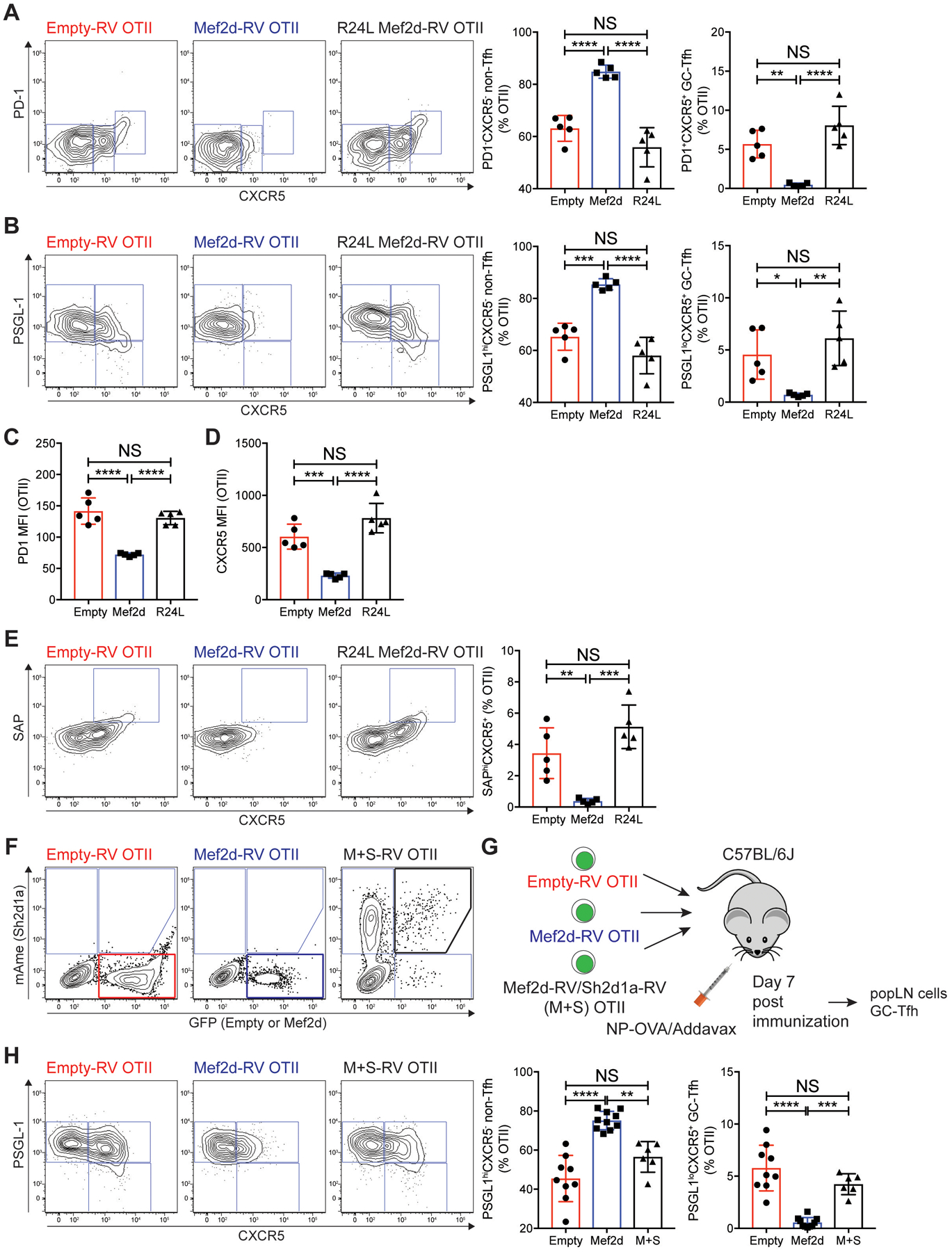
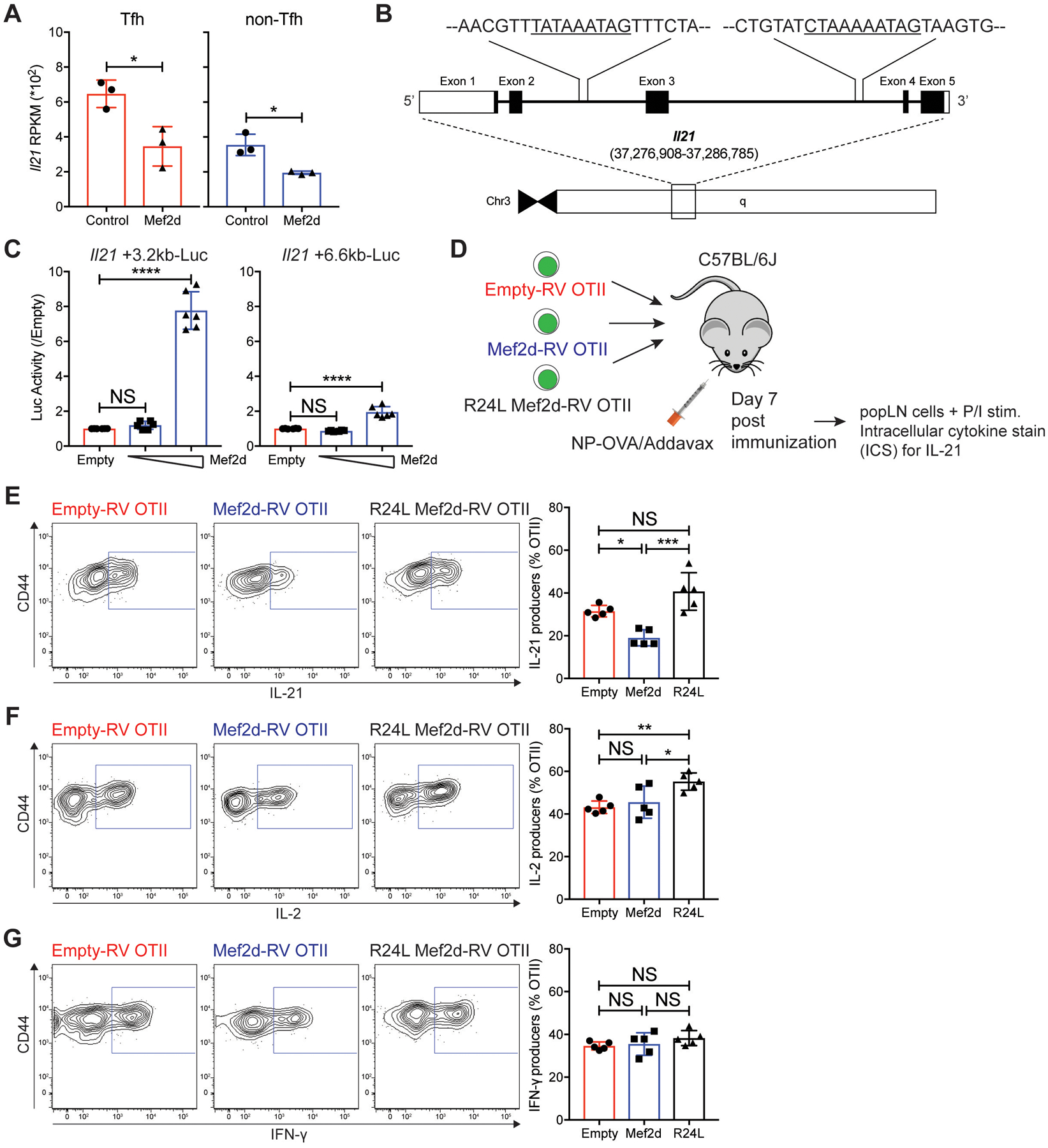
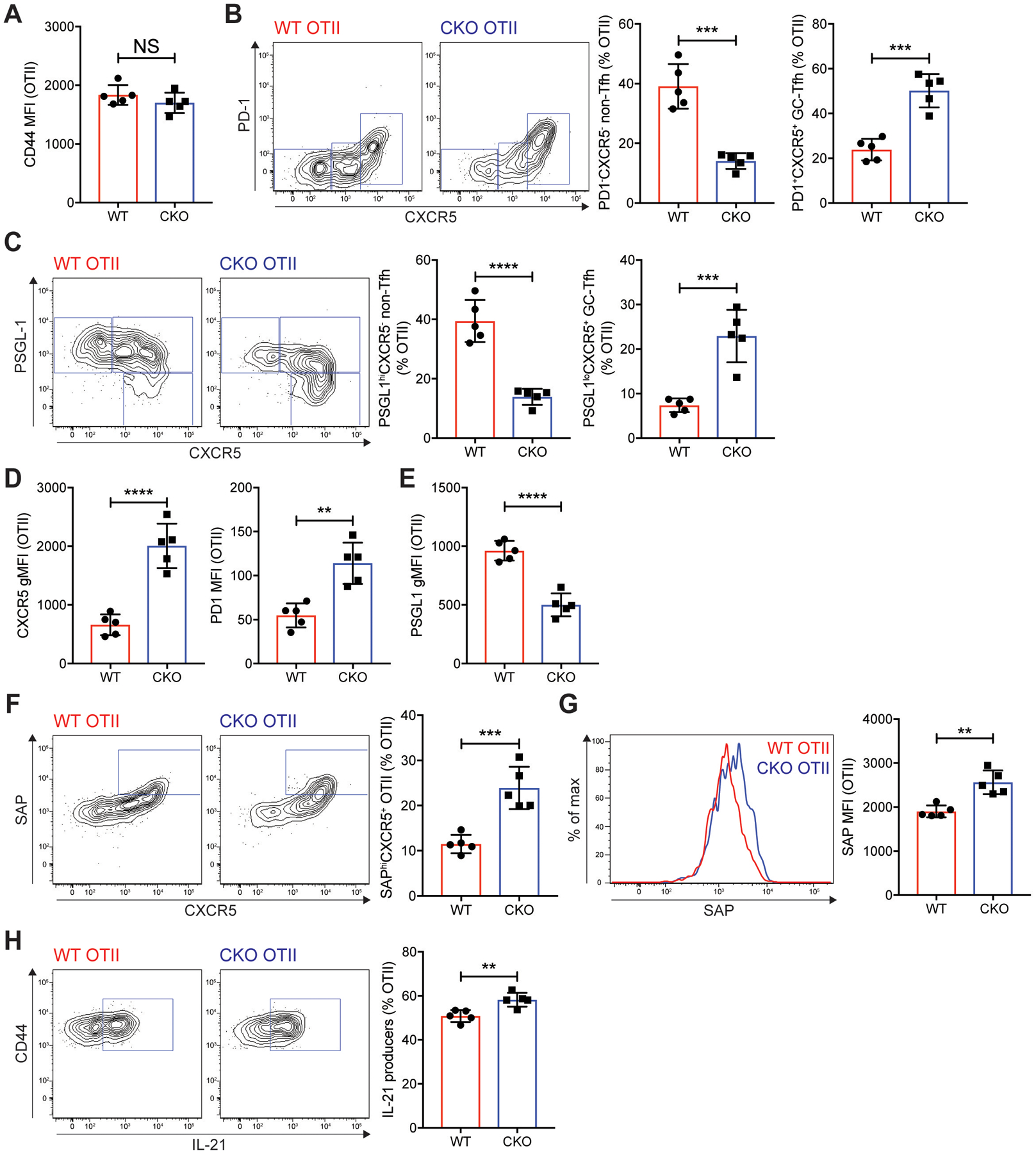
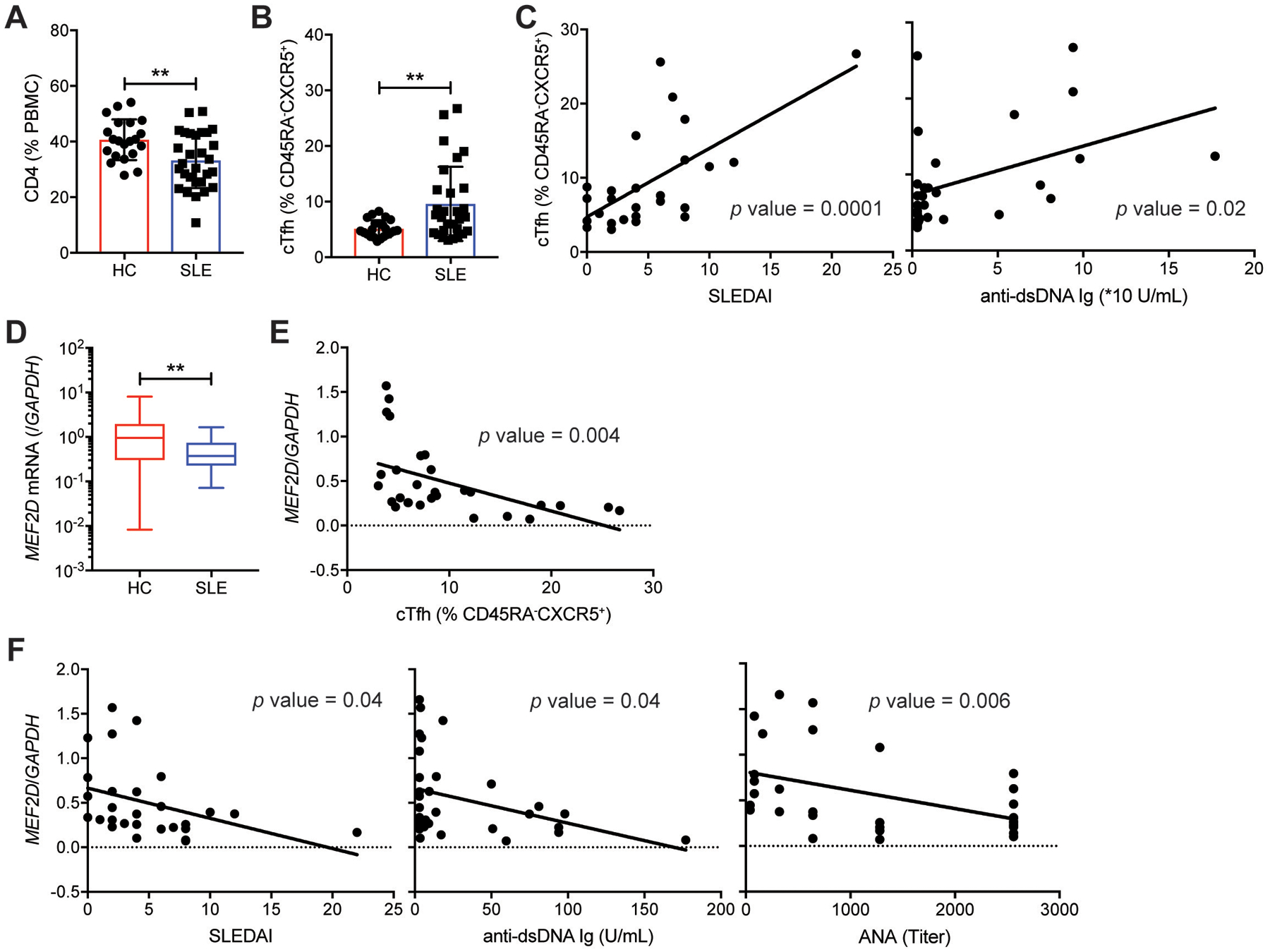
Communication between CD4 T cells and cognate B cells is key for the former to fully mature into germinal center-T follicular helper (GC-TFH) cells and for the latter to mount a CD4 T cell-dependent humoral immune response. Although this interaction occurs in a B:T synapse-dependent manner, how CD4 T cells transcriptionally regulate B:T synapse formation remains largely unknown. Here, we report that Mef2d, an isoform of the myocyte enhancer factor 2 (Mef2) transcription factor family, is a critical regulator of this process. In CD4 T cells, Mef2d negatively regulates expression of Sh2d1a, which encodes SLAM-associated protein (SAP), a critical regulator of B:T synapses. We found that Mef2d regulates Sh2d1a expression via DNA binding-dependent transcriptional repression, inhibiting SAP-dependent B:T synapse formation and preventing antigen-specific CD4 T cells from differentiating into GC-TFH cells. Mef2d also impeded IL-21 production by CD4 T cells, an important B cell help signaling molecule, via direct repression of the Il21 gene. In contrast, CD4 T cell-specific disruption of Mef2d led to a substantial increase in GC-TFH differentiation in response to protein immunization, concurrent with enhanced SAP expression. MEF2D mRNA expression inversely correlates with human systemic lupus erythematosus (SLE) patient autoimmune parameters, including circulating TFH-like cell frequencies, autoantibodies, and SLEDAI scores. These findings highlight Mef2d as a pivotal rheostat in CD4 T cells for controlling GC formation and antibody production by B cells.
Conflict of interest statement
Figures
References
-
- Dustin ML, Colman DR, Neural and immunological synaptic relations. Science 298, 785–789 (2002). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
