

Structural constraints link differences in neutralization potency of human anti-Eastern equine encephalitis virus monoclonal antibodies
- PMID: 36961925
- PMCID: PMC10068833
- DOI: 10.1073/pnas.2213690120
Structural constraints link differences in neutralization potency of human anti-Eastern equine encephalitis virus monoclonal antibodies
Abstract
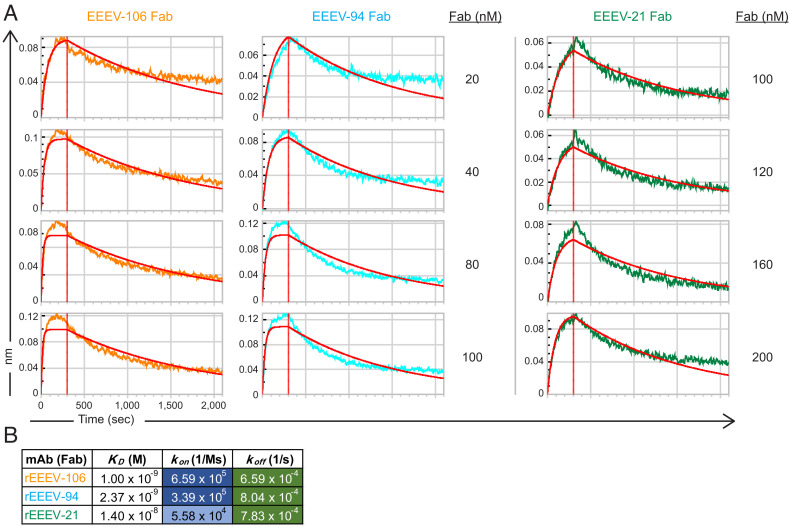
Selection and development of monoclonal antibody (mAb) therapeutics against pathogenic viruses depends on certain functional characteristics. Neutralization potency, or the half-maximal inhibitory concentration (IC50) values, is an important characteristic of candidate therapeutic antibodies. Structural insights into the bases of neutralization potency differences between antiviral neutralizing mAbs are lacking. In this report, we present cryo-electron microscopy (EM) reconstructions of three anti-Eastern equine encephalitis virus (EEEV) neutralizing human mAbs targeting overlapping epitopes on the E2 protein, with greater than 20-fold differences in their respective IC50 values. From our structural and biophysical analyses, we identify several constraints that contribute to the observed differences in the neutralization potencies. Cryo-EM reconstructions of EEEV in complex with these Fab fragments reveal structural constraints that dictate intravirion or intervirion cross-linking of glycoprotein spikes by their IgG counterparts as a mechanism of neutralization. Additionally, we describe critical features for the recognition of EEEV by these mAbs including the epitope-paratope interaction surface, occupancy, and kinetic differences in on-rate for binding to the E2 protein. Each constraint contributes to the extent of EEEV inhibition for blockade of virus entry, fusion, and/or egress. These findings provide structural and biophysical insights into the differences in mechanism and neutralization potencies of these antibodies, which help inform rational design principles for candidate vaccines and therapeutic antibodies for all icosahedral viruses.
Keywords: alphavirus; antibodies, human monoclonal; cryo-EM; neutralization; therapy.
Conflict of interest statement
L.E.W. serves as a consultant for BigHat Biosciences. The content of this article is solely the responsibility of the authors and does not represent the official views of BigHat Biosciences. J.E.C. has served as a consultant for Luna Labs USA, Merck Sharp & Dohme Corporation, Emergent Biosolutions, GlaxoSmithKline, and BTG International Inc, is a member of the Scientific Advisory Board of Meissa Vaccines, a former member of the Scientific Advisory Board of Gigagen (Grifols), and is founder of IDBiologics. The laboratory of J.E.C. received unrelated sponsored research agreements from AstraZeneca, Takeda, and IDBiologics during the conduct of the study. The Crowe laboratory at Vanderbilt University Medical Center has received unrelated sponsored research agreements from IDBiologics, Takeda Pharmaceuticals, and AstraZeneca. All other authors report no conflicts. J.E.C. has owns stock in IDBiologics and has stock options in Meissa Vaccines. Vanderbilt University has applied for a patent related to antibodies described in this paper. The Crowe laboratory at Vanderbilt University Medical Center has received unrelated sponsored research agreements from IDBiologics, Takeda Pharmaceuticals, and AstraZeneca. J.E.C. is a member of the Scientific Advisory Board of Meissa Vaccines, a former member of the Gigagen Scientific Advisory Board, and is Founder of IDBiologics.
Figures
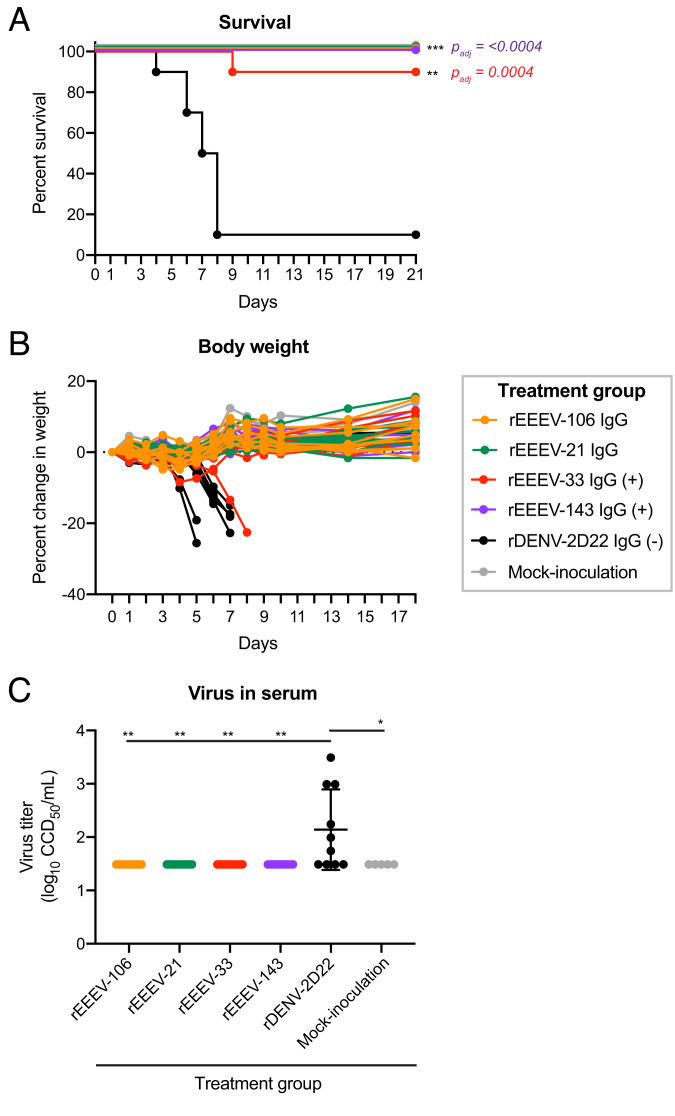

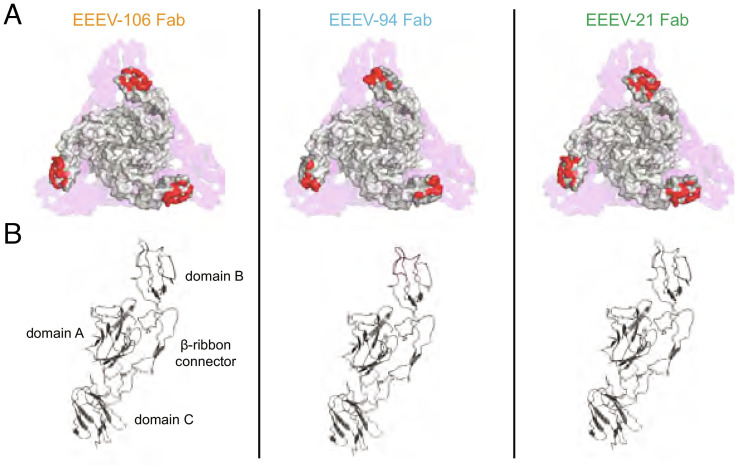


Similar articles
-
Structural elucidation of a unique binding mode by an intact alphavirus human IgG molecule to a quaternary epitope.Nat Commun. 2025 Aug 19;16(1):7716. doi: 10.1038/s41467-025-60505-x. Nat Commun. 2025. PMID: 40830099 Free PMC article.
-
Human Antibodies Protect against Aerosolized Eastern Equine Encephalitis Virus Infection.Cell. 2020 Dec 23;183(7):1884-1900.e23. doi: 10.1016/j.cell.2020.11.011. Epub 2020 Dec 9. Cell. 2020. PMID: 33301709 Free PMC article.
-
Cryo-EM Structures of Eastern Equine Encephalitis Virus Reveal Mechanisms of Virus Disassembly and Antibody Neutralization.Cell Rep. 2018 Dec 11;25(11):3136-3147.e5. doi: 10.1016/j.celrep.2018.11.067. Cell Rep. 2018. PMID: 30540945 Free PMC article.
-
Protective antibodies against Eastern equine encephalitis virus bind to epitopes in domains A and B of the E2 glycoprotein.Nat Microbiol. 2019 Jan;4(1):187-197. doi: 10.1038/s41564-018-0286-4. Epub 2018 Nov 19. Nat Microbiol. 2019. PMID: 30455470 Free PMC article.
-
Eastern Equine Encephalitis Virus: A Case Report and Brief Literature Review of Current Therapeutic and Preventative Strategies.Vector Borne Zoonotic Dis. 2024 Feb;24(2):118-121. doi: 10.1089/vbz.2023.0011. Epub 2023 Oct 23. Vector Borne Zoonotic Dis. 2024. PMID: 37870590 Review.
Cited by
-
Molecular mechanism and structure-guided humanization of a broadly neutralizing antibody against SFTSV.PLoS Pathog. 2024 Sep 25;20(9):e1012550. doi: 10.1371/journal.ppat.1012550. eCollection 2024 Sep. PLoS Pathog. 2024. PMID: 39321193 Free PMC article.
-
Generating prophylactic immunity against arboviruses in vertebrates and invertebrates.Nat Rev Immunol. 2024 Sep;24(9):621-636. doi: 10.1038/s41577-024-01016-6. Epub 2024 Apr 3. Nat Rev Immunol. 2024. PMID: 38570719 Review.
-
Insights into the Landscape of Alphavirus Receptor and Antibody Interactions.Viruses. 2025 Jul 21;17(7):1019. doi: 10.3390/v17071019. Viruses. 2025. PMID: 40733635 Free PMC article. Review.
-
Structural elucidation of a unique binding mode by an intact alphavirus human IgG molecule to a quaternary epitope.Nat Commun. 2025 Aug 19;16(1):7716. doi: 10.1038/s41467-025-60505-x. Nat Commun. 2025. PMID: 40830099 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
