

Experimental Tests of the Virtual Circular Genome Model for Nonenzymatic RNA Replication
- PMID: 36963403
- PMCID: PMC10080680
- DOI: 10.1021/jacs.3c00612
Experimental Tests of the Virtual Circular Genome Model for Nonenzymatic RNA Replication
Abstract
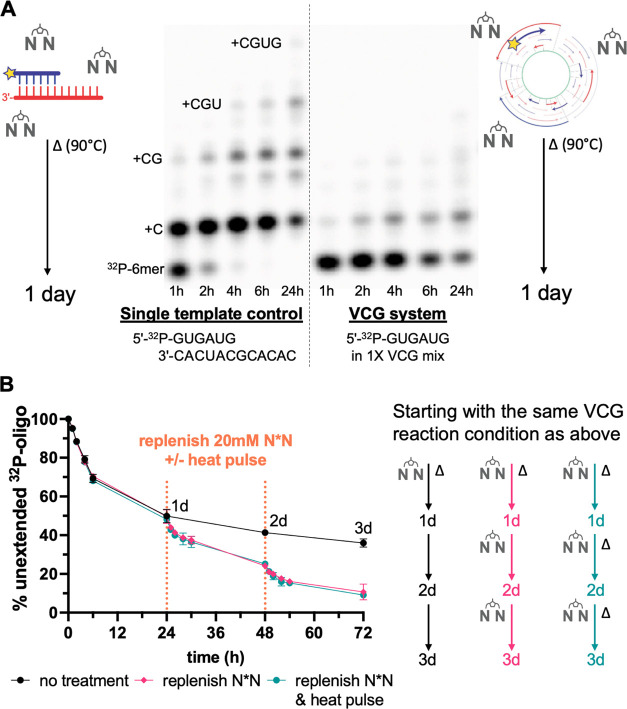
The virtual circular genome (VCG) model was proposed as a means of going beyond template copying to indefinite cycles of nonenzymatic RNA replication during the origin of life. In the VCG model, the protocellular genome is a collection of short oligonucleotides that map to both strands of a virtual circular sequence. Replication is driven by templated nonenzymatic primer extensions on a subset of kinetically trapped partially base-paired configurations, followed by the shuffling of these configurations to enable continued oligonucleotide elongation. Here, we describe initial experimental studies of the feasibility of the VCG model for replication. We designed a small 12-nucleotide model VCG and synthesized all 247 oligonucleotides of lengths 2 to 12 corresponding to this genome. We experimentally monitored the fate of individual labeled primers in the pool of VCG oligonucleotides following the addition of activated nucleotides and investigated the effect of factors such as oligonucleotide length, concentration, composition, and temperature on the extent of primer extension. We observe a surprisingly prolonged equilibration process in the VCG system that enables a considerable extent of reaction. We find that environmental fluctuations would be essential for continuous templated extension of the entire VCG system since the shortest oligonucleotides can only bind to templates at low temperatures, while the longest oligonucleotides require high-temperature spikes to escape from inactive configurations. Finally, we demonstrate that primer extension is significantly enhanced when the mix of VCG oligonucleotides is preactivated. We discuss the necessity of ongoing in situ activation chemistry for continuous and accurate VCG replication.
Conflict of interest statement
The authors declare no competing financial interest.
Figures





Similar articles
-
Enhanced nonenzymatic RNA copying with in-situ activation of short oligonucleotides.Nucleic Acids Res. 2023 Jul 21;51(13):6528-6539. doi: 10.1093/nar/gkad439. Nucleic Acids Res. 2023. PMID: 37247941 Free PMC article.
-
Overcoming nucleotide bias in the nonenzymatic copying of RNA templates.Nucleic Acids Res. 2024 Dec 11;52(22):13515-13529. doi: 10.1093/nar/gkae982. Nucleic Acids Res. 2024. PMID: 39530216 Free PMC article.
-
The virtual circular genome model for primordial RNA replication.RNA. 2021 Jan;27(1):1-11. doi: 10.1261/rna.077693.120. Epub 2020 Oct 7. RNA. 2021. PMID: 33028653 Free PMC article.
-
The Mechanism of Nonenzymatic Template Copying with Imidazole-Activated Nucleotides.Angew Chem Int Ed Engl. 2019 Aug 5;58(32):10812-10819. doi: 10.1002/anie.201902050. Epub 2019 Jul 4. Angew Chem Int Ed Engl. 2019. PMID: 30908802 Review.
-
The Emergence of RNA from the Heterogeneous Products of Prebiotic Nucleotide Synthesis.J Am Chem Soc. 2021 Mar 10;143(9):3267-3279. doi: 10.1021/jacs.0c12955. Epub 2021 Feb 26. J Am Chem Soc. 2021. PMID: 33636080 Review.
Cited by
-
Enhanced nonenzymatic RNA copying with in-situ activation of short oligonucleotides.Nucleic Acids Res. 2023 Jul 21;51(13):6528-6539. doi: 10.1093/nar/gkad439. Nucleic Acids Res. 2023. PMID: 37247941 Free PMC article.
-
Overcoming nucleotide bias in the nonenzymatic copying of RNA templates.Nucleic Acids Res. 2024 Dec 11;52(22):13515-13529. doi: 10.1093/nar/gkae982. Nucleic Acids Res. 2024. PMID: 39530216 Free PMC article.
-
Nonenzymatic RNA copying with a potentially primordial genetic alphabet.Proc Natl Acad Sci U S A. 2025 May 27;122(21):e2505720122. doi: 10.1073/pnas.2505720122. Epub 2025 May 21. Proc Natl Acad Sci U S A. 2025. PMID: 40397670 Free PMC article.
-
Polymerization and replication of primordial RNA induced by clay-water interface dynamics.Commun Chem. 2025 Aug 7;8(1):236. doi: 10.1038/s42004-025-01632-w. Commun Chem. 2025. PMID: 40775023 Free PMC article.
-
Highlights from the 57th Bürgenstock Conference on Stereochemistry 2024.Chem Sci. 2024 Jun 13;15(25):9392-9396. doi: 10.1039/d4sc90102a. eCollection 2024 Jun 26. Chem Sci. 2024. PMID: 38939160 Free PMC article.
References
-
- Szostak J. W. The Eightfold Path to Non-Enzymatic RNA Replication. J. Syst. Chem. 2012, 3, 210.1186/1759-2208-3-2. - DOI
-
- Martin C.; Frenkel-Pinter M.; Smith K. H.; Rivera-Santana V. F.; Sargon A. B.; Jacobson K. C.; Guzman-Martinez A.; Williams L. D.; Leman L. J.; Liotta C. L.; Grover M. A.; Hud N. V. Water-Based Dynamic Depsipeptide Chemistry: Building Block Recycling and Oligomer Distribution Control Using Hydration–Dehydration Cycles. JACS Au 2022, 2, 1395–1404. 10.1021/jacsau.2c00087. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
