

The Batten disease protein CLN3 is important for stress granules dynamics and translational activity
- PMID: 36965618
- PMCID: PMC10149212
- DOI: 10.1016/j.jbc.2023.104649
The Batten disease protein CLN3 is important for stress granules dynamics and translational activity
Abstract
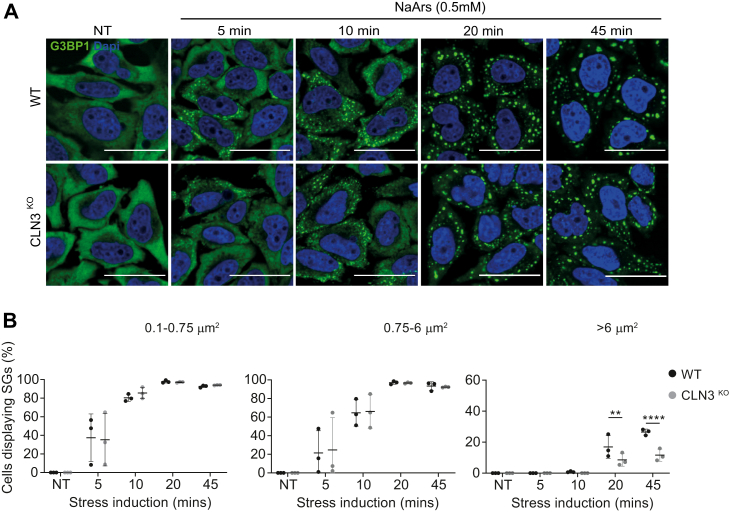
The assembly of membrane-less organelles such as stress granules (SGs) is emerging as central in helping cells rapidly respond and adapt to stress. Following stress sensing, the resulting global translational shutoff leads to the condensation of stalled mRNAs and proteins into SGs. By reorganizing cytoplasmic contents, SGs can modulate RNA translation, biochemical reactions, and signaling cascades to promote survival until the stress is resolved. While mechanisms for SG disassembly are not widely understood, the resolution of SGs is important for maintaining cell viability and protein homeostasis. Mutations that lead to persistent or aberrant SGs are increasingly associated with neuropathology and a hallmark of several neurodegenerative diseases. Mutations in CLN3 are causative of juvenile neuronal ceroid lipofuscinosis, a rare neurodegenerative disease affecting children also known as Batten disease. CLN3 encodes a transmembrane lysosomal protein implicated in autophagy, endosomal trafficking, metabolism, and response to oxidative stress. Using a HeLa cell model lacking CLN3, we now show that CLN3KO is associated with an altered metabolic profile, reduced global translation, and altered stress signaling. Furthermore, loss of CLN3 function results in perturbations in SG dynamics, resulting in assembly and disassembly defects, and altered expression of the key SG nucleating factor G3BP1. With a growing interest in SG-modulating drugs for the treatment of neurodegenerative diseases, novel insights into the molecular basis of CLN3 Batten disease may reveal avenues for disease-modifying treatments for this debilitating childhood disease.
Keywords: Batten disease; CLN3; NCL; metabolism; stress; stress granules; translation control.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures








References
-
- Williams R.E., Mole S.E. New nomenclature and classification scheme for the neuronal ceroid lipofuscinoses. Neurology. 2012;79:183–191. - PubMed
-
- Kitzmuller C., Haines R.L., Codlin S., Cutler D.F., Mole S.E. A function retained by the common mutant CLN3 protein is responsible for the late onset of juvenile neuronal ceroid lipofuscinosis. Hum. Mol. Genet. 2008;17:303–312. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BB/V014528/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- MR/S022465/1/MRC_/Medical Research Council/United Kingdom
- MR/R02426×/1/MRC_/Medical Research Council/United Kingdom
- DH_/Department of Health/United Kingdom
- MR/R02426X/1/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
