

Regulation of nucleotide metabolism in cancers and immune disorders
- PMID: 36967301
- PMCID: PMC10518033
- DOI: 10.1016/j.tcb.2023.03.003
Regulation of nucleotide metabolism in cancers and immune disorders
Abstract
Nucleotides are the foundational elements of life. Proliferative cells acquire nutrients for energy production and the synthesis of macromolecules, including proteins, lipids, and nucleic acids. Nucleotides are continuously replenished through the activation of the nucleotide synthesis pathways. Despite the importance of nucleotides in cell physiology, there is still much to learn about how the purine and pyrimidine synthesis pathways are regulated in response to intracellular and exogenous signals. Over the past decade, evidence has emerged that several signaling pathways [Akt, mechanistic target of rapamycin complex I (mTORC1), RAS, TP53, and Hippo-Yes-associated protein (YAP) signaling] alter nucleotide synthesis activity and influence cell function. Here, we examine the mechanisms by which these signaling networks affect de novo nucleotide synthesis in mammalian cells. We also discuss how these molecular links can be targeted in diseases such as cancers and immune disorders.
Keywords: cancer metabolism; de novo purine and pyrimidine synthesis; immune disorders; metabolic vulnerability; nucleotide signaling; signaling pathways.
Copyright © 2023 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
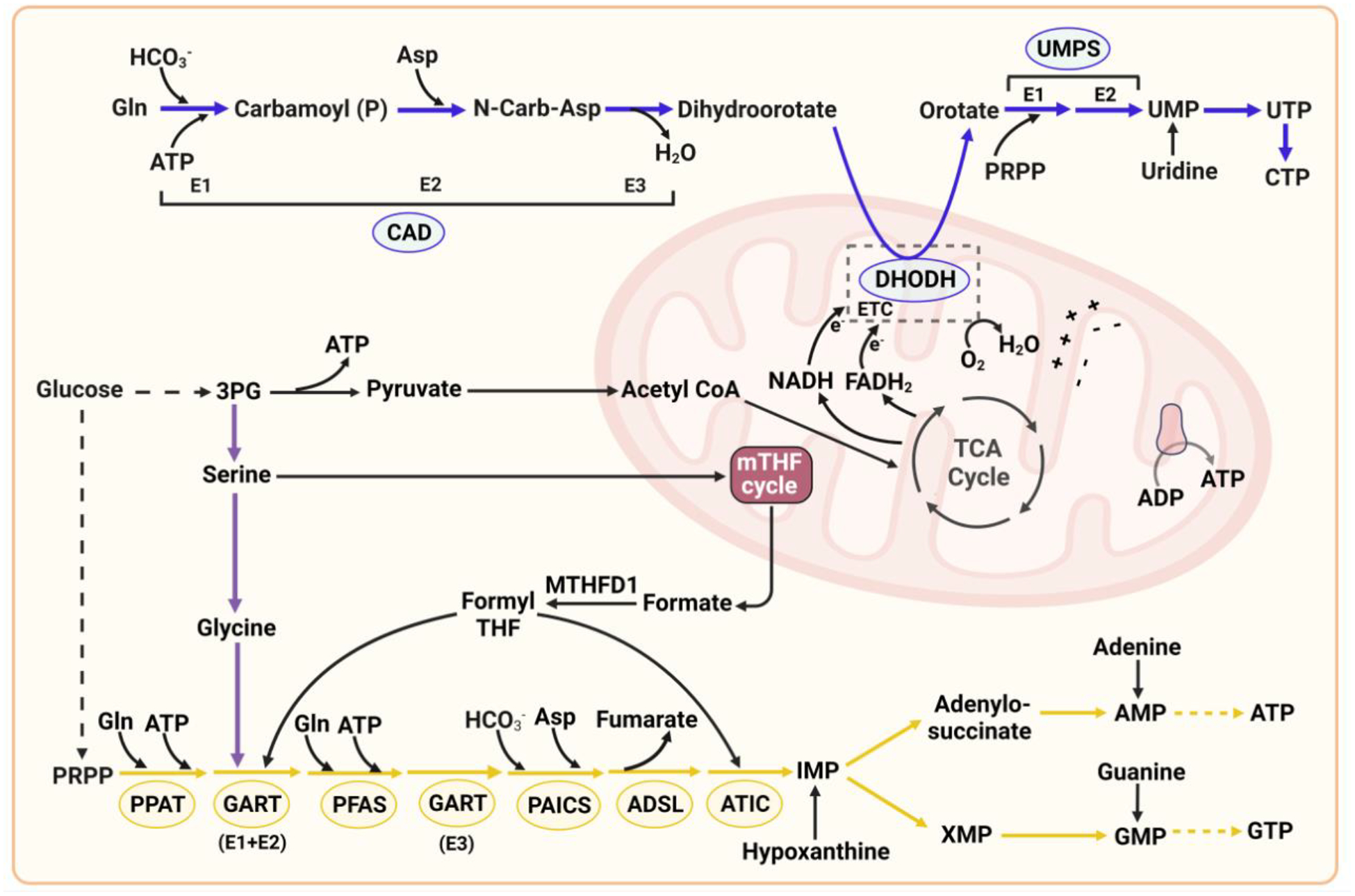
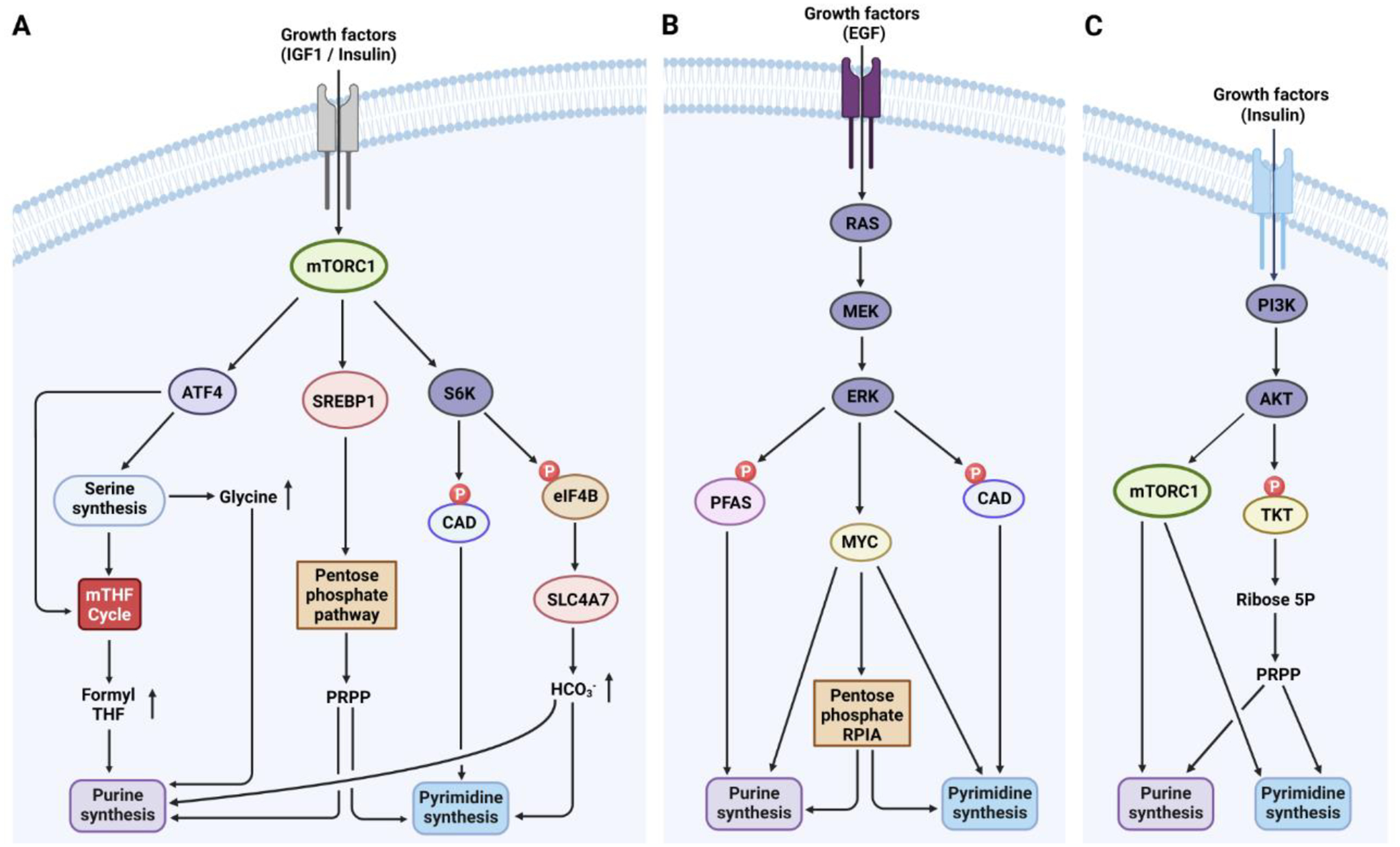
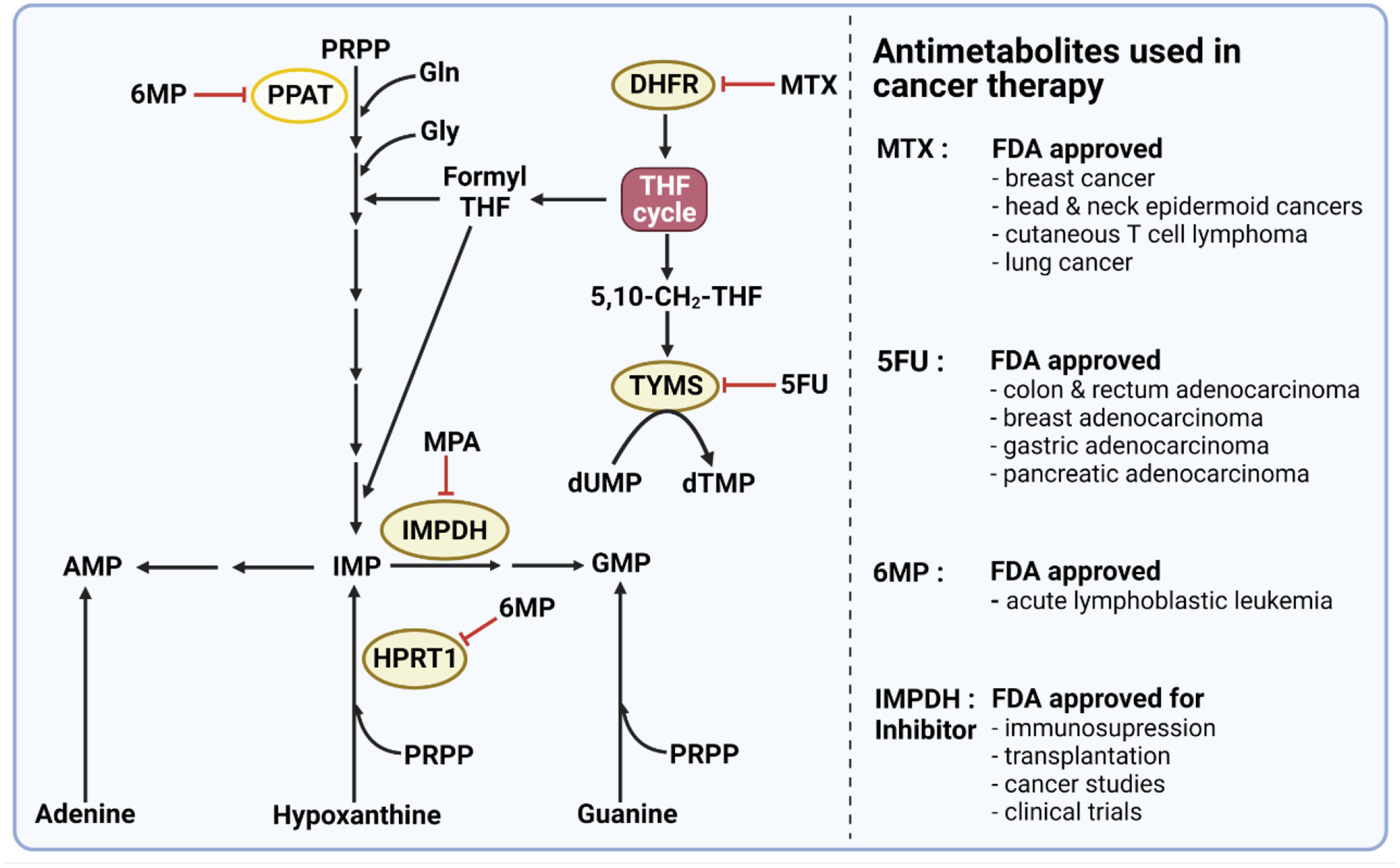
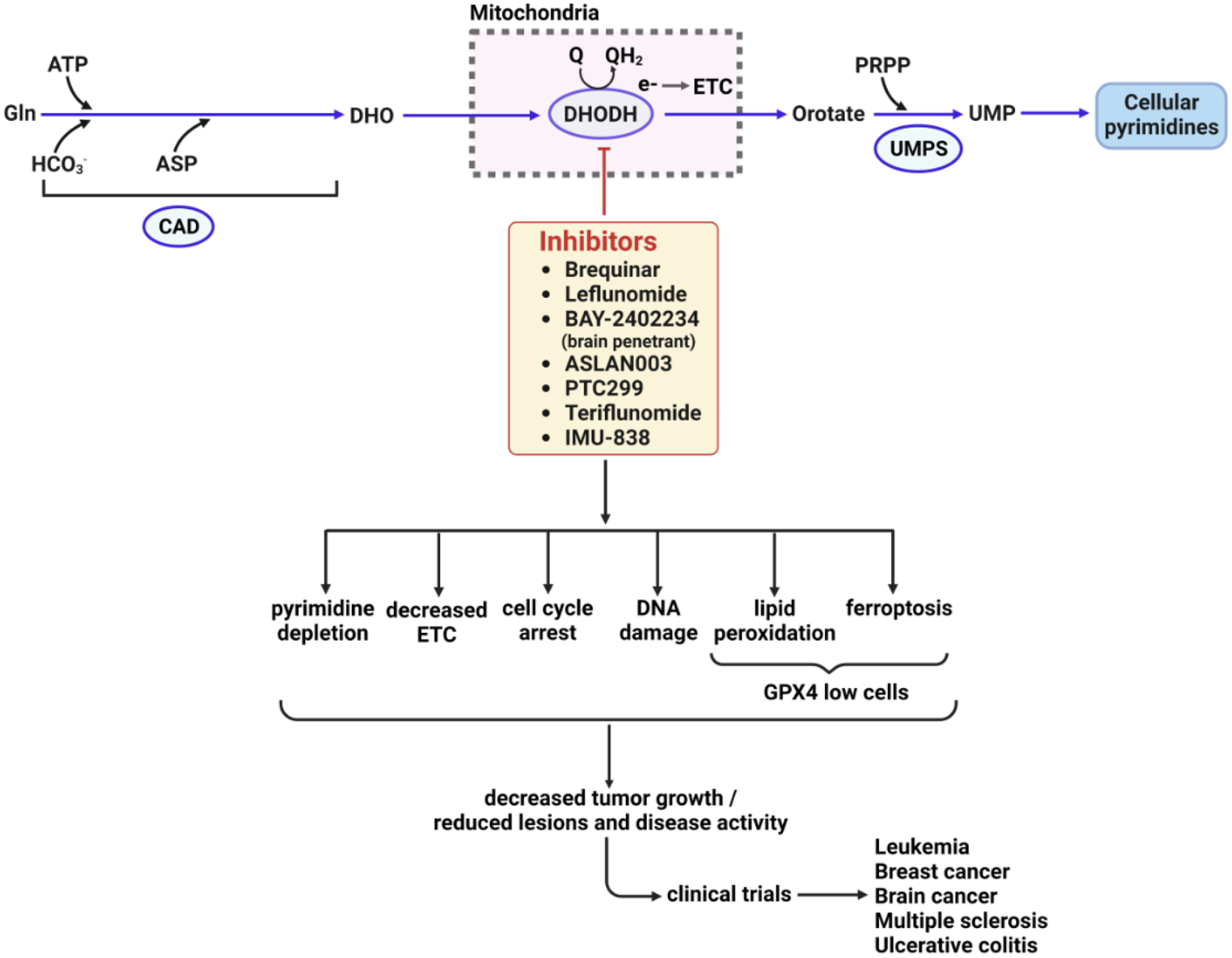
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous