

Quantification and Mapping of Alkylation in the Human Genome Reveal Single Nucleotide Resolution Precursors of Mutational Signatures
- PMID: 36968528
- PMCID: PMC10037492
- DOI: 10.1021/acscentsci.2c01100
Quantification and Mapping of Alkylation in the Human Genome Reveal Single Nucleotide Resolution Precursors of Mutational Signatures
Erratum in
-
Correction to "Quantification and Mapping of Alkylation in the Human Genome Reveal Single Nucleotide Resolution Precursors of Mutational Signatures".ACS Cent Sci. 2024 Jan 25;10(2):487. doi: 10.1021/acscentsci.3c01597. eCollection 2024 Feb 28. ACS Cent Sci. 2024. PMID: 38435521 Free PMC article.
Abstract
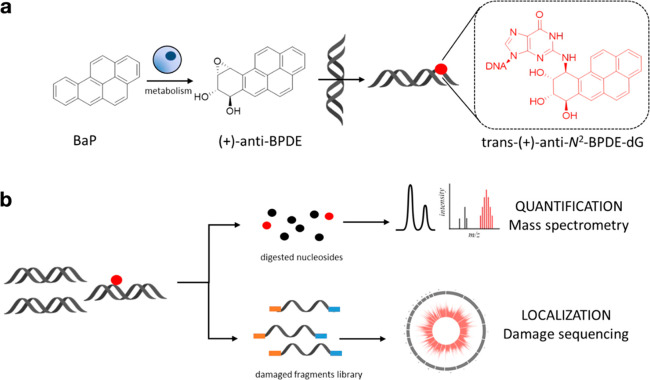
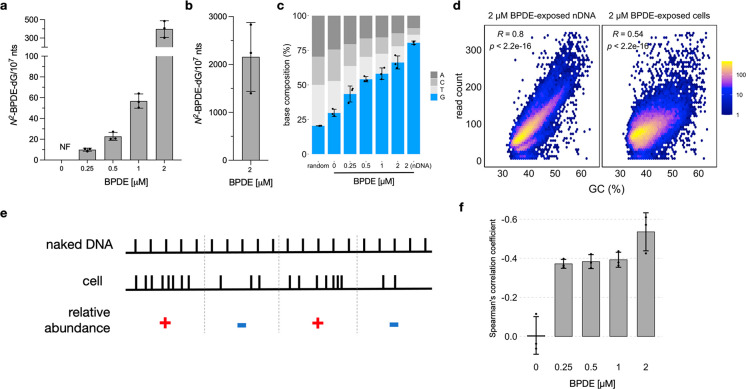
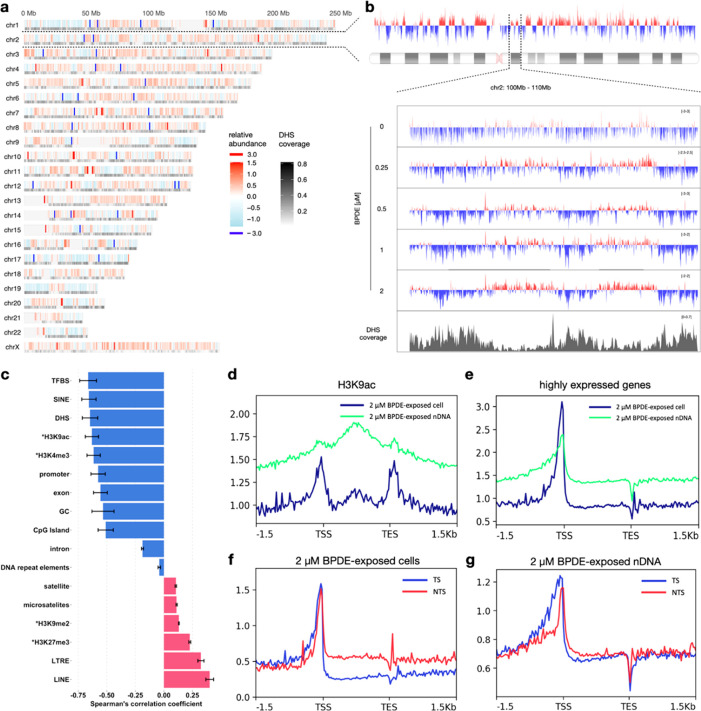
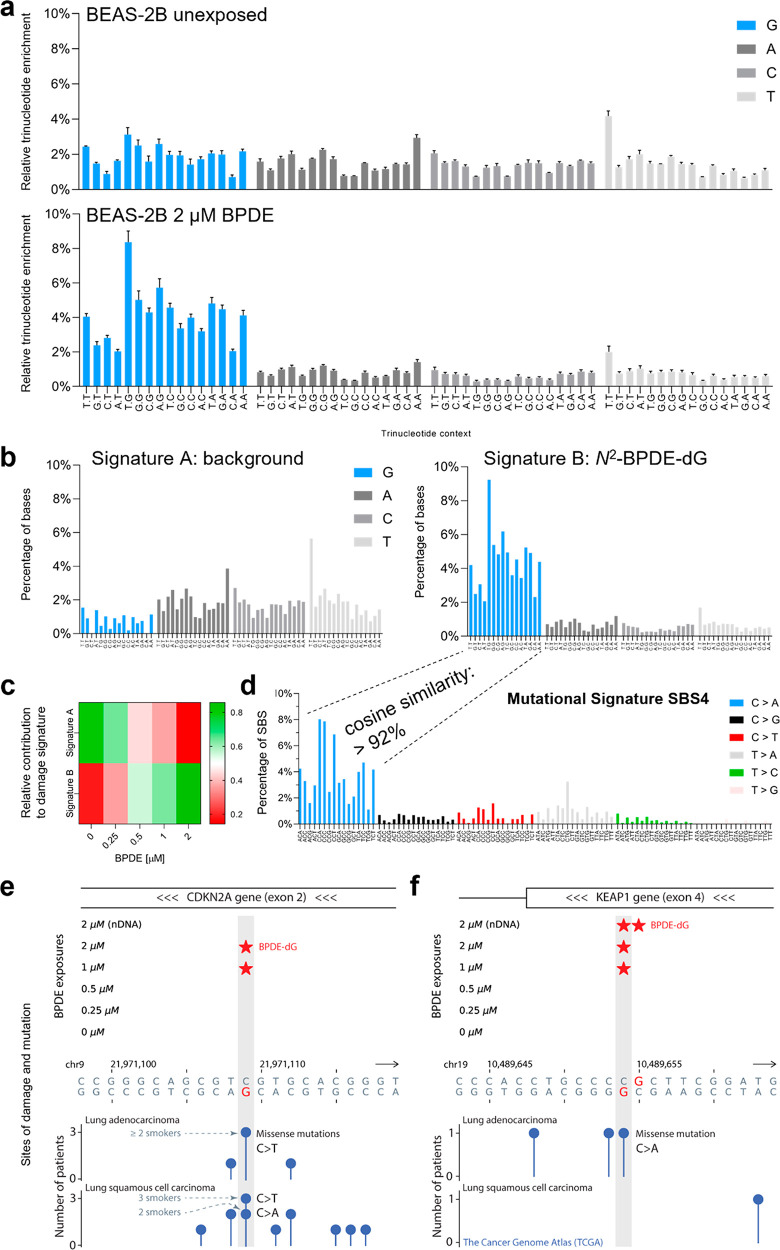
Chemical modifications to DNA bases, including DNA adducts arising from reactions with electrophilic chemicals, are well-known to impact cell growth, miscode during replication, and influence disease etiology. However, knowledge of how genomic sequences and structures influence the accumulation of alkylated DNA bases is not broadly characterized with high resolution, nor have these patterns been linked with overall quantities of modified bases in the genome. For benzo(a) pyrene (BaP), a ubiquitous environmental carcinogen, we developed a single-nucleotide resolution damage sequencing method to map in a human lung cell line the main mutagenic adduct arising from BaP. Furthermore, we combined this analysis with quantitative mass spectrometry to evaluate the dose-response profile of adduct formation. By comparing damage abundance with DNase hypersensitive sites, transcription levels, and other genome annotation data, we found that although overall adduct levels rose with increasing chemical exposure concentration, genomic distribution patterns consistently correlated with chromatin state and transcriptional status. Moreover, due to the single nucleotide resolution characteristics of this DNA damage map, we could determine preferred DNA triad sequence contexts for alkylation accumulation, revealing a characteristic DNA damage signature. This new BaP damage signature had a profile highly similar to mutational signatures identified previously in lung cancer genomes from smokers. Thus, these data provide insight on how genomic features shape the accumulation of alkylation products in the genome and predictive strategies for linking single-nucleotide resolution in vitro damage maps with human cancer mutations.
© 2023 The Authors. Published by American Chemical Society.
Conflict of interest statement
The authors declare the following competing financial interest(s): J.H., N.S., and N.V.I. are employees of Philip Morris International.
Figures
References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
