

ROS-removing nano-medicine for navigating inflammatory microenvironment to enhance anti-epileptic therapy
- PMID: 36970212
- PMCID: PMC10031259
- DOI: 10.1016/j.apsb.2022.09.019
ROS-removing nano-medicine for navigating inflammatory microenvironment to enhance anti-epileptic therapy
Abstract
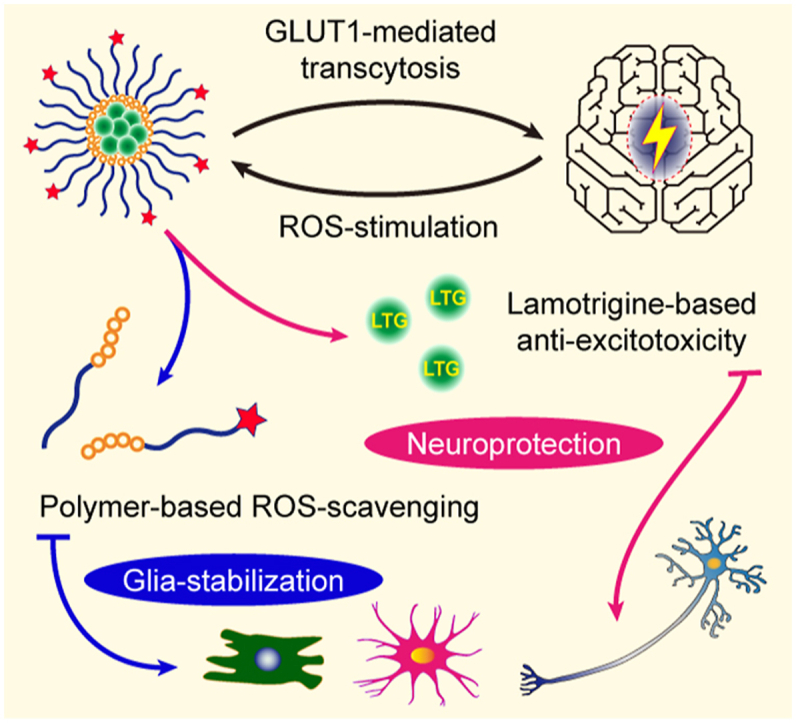
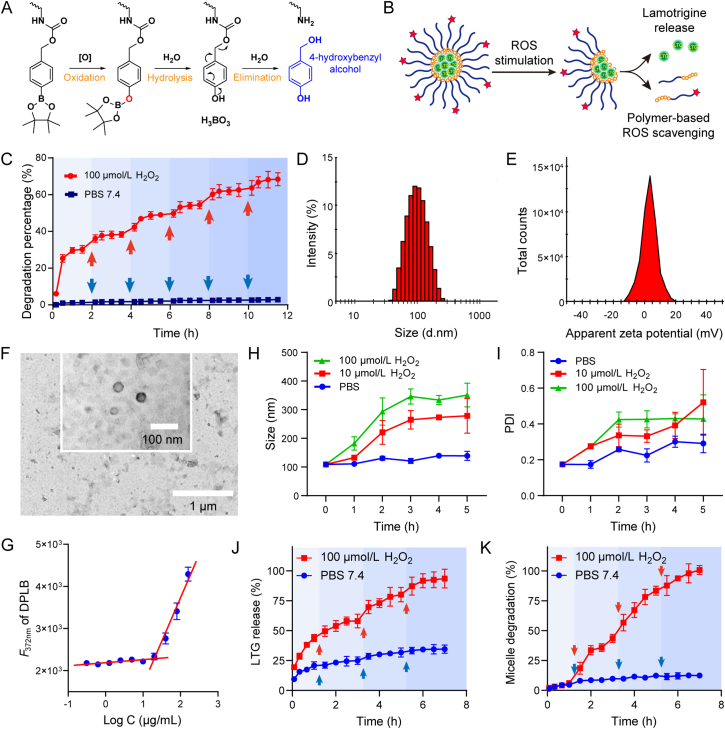
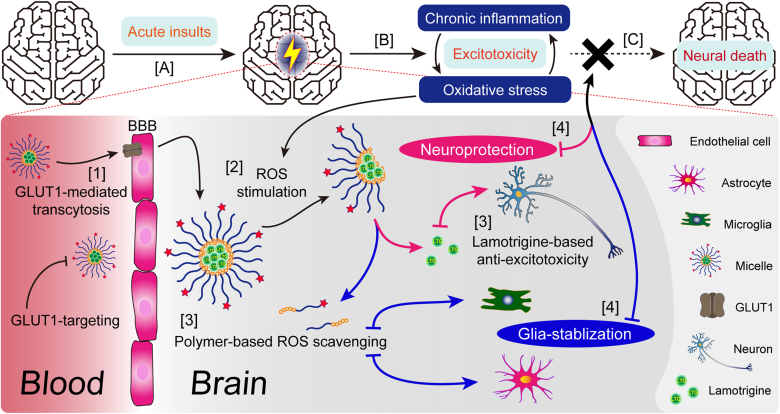
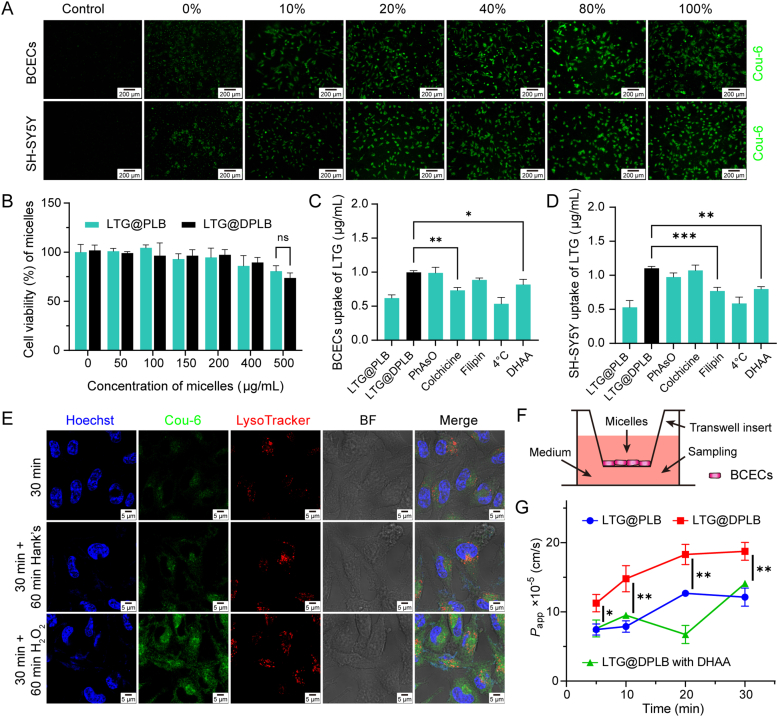
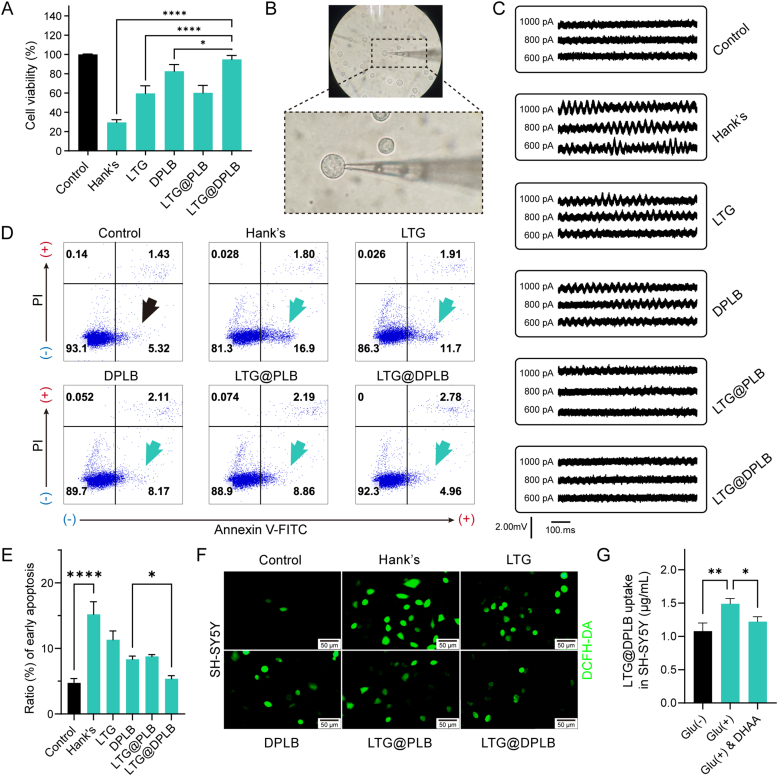
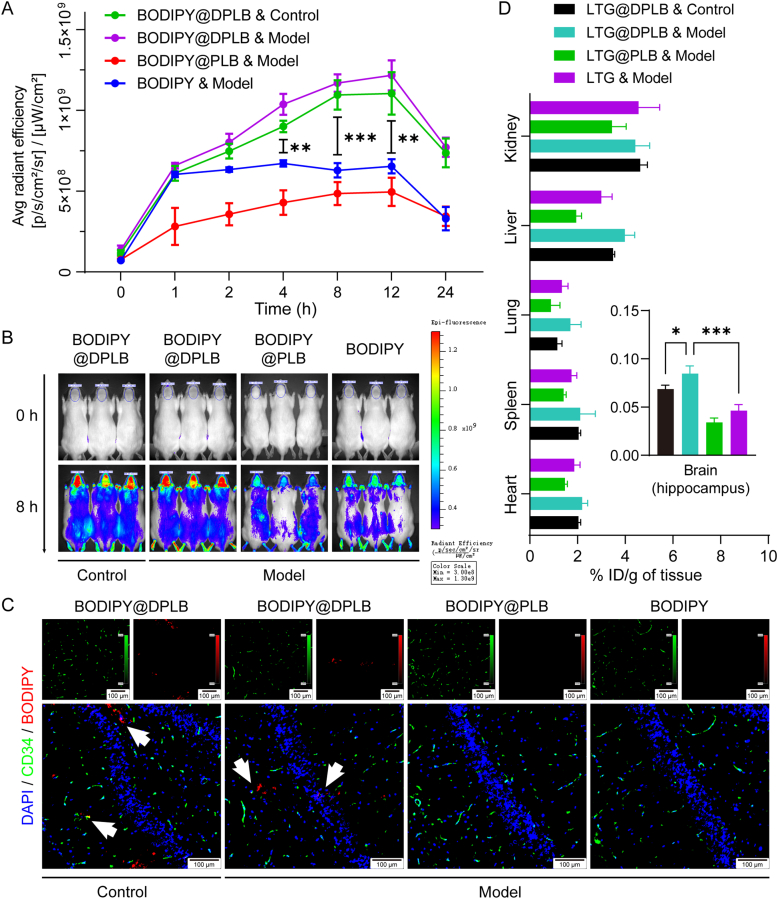
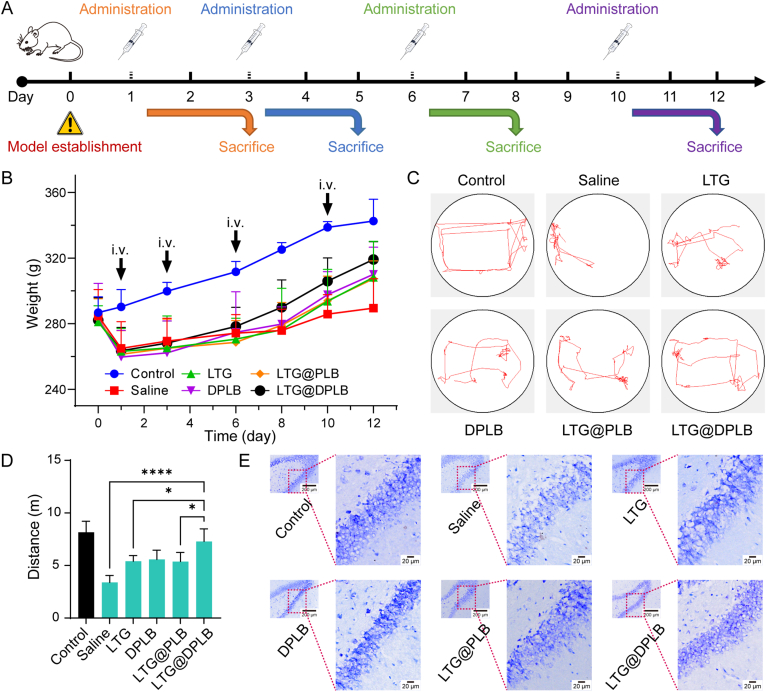
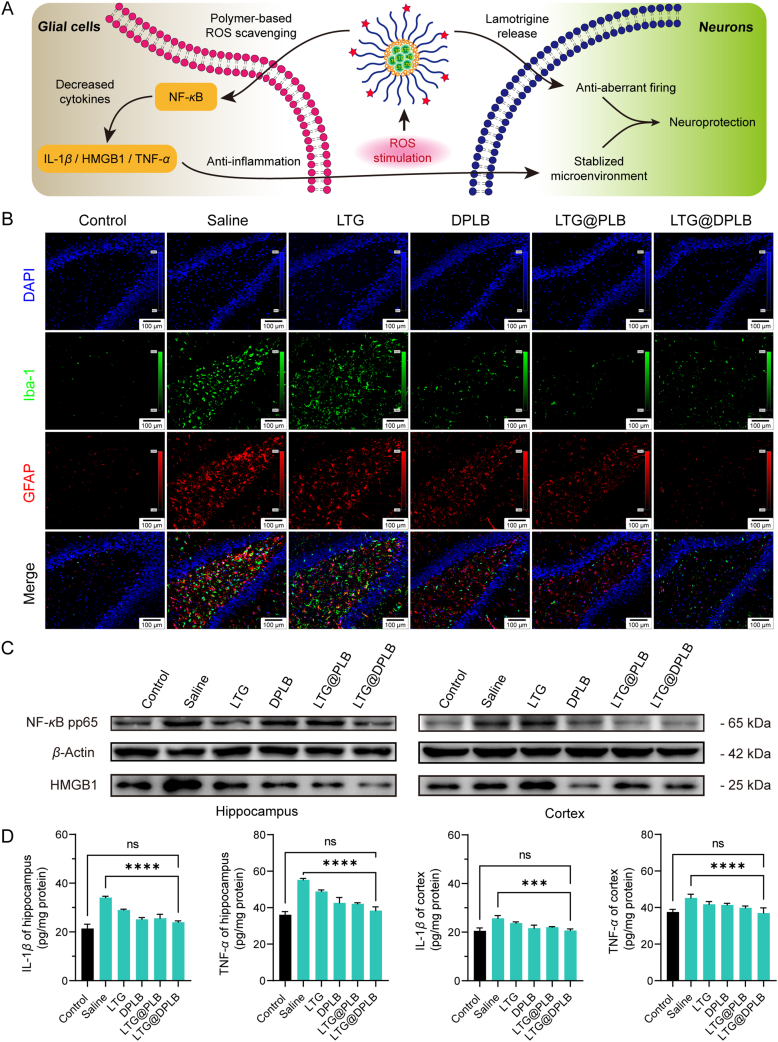
As a neurological disorder in the brain, epilepsy is not only associated with abnormal synchronized discharging of neurons, but also inseparable from non-neuronal elements in the altered microenvironment. Anti-epileptic drugs (AEDs) merely focusing on neuronal circuits frequently turn out deficient, which is necessitating comprehensive strategies of medications to cover over-exciting neurons, activated glial cells, oxidative stress and chronic inflammation synchronously. Therefore, we would report the design of a polymeric micelle drug delivery system that was functioned with brain targeting and cerebral microenvironment modulation. In brief, reactive oxygen species (ROS)-sensitive phenylboronic ester was conjugated with poly-ethylene glycol (PEG) to form amphiphilic copolymers. Additionally, dehydroascorbic acid (DHAA), an analogue of glucose, was applied to target glucose transporter 1 (GLUT1) and facilitate micelle penetration across the blood‒brain barrier (BBB). A classic hydrophobic AED, lamotrigine (LTG), was encapsulated in the micelles via self-assembly. When administrated and transferred across the BBB, ROS-scavenging polymers were expected to integrate anti-oxidation, anti-inflammation and neuro-electric modulation into one strategy. Moreover, micelles would alter LTG distribution in vivo with improved efficacy. Overall, the combined anti-epileptic therapy might provide effective opinions on how to maximize neuroprotection during early epileptogenesis.
Keywords: Epilepsy; Gliosis; Inflammation; Neuroprotection; Polymeric micelle; Reactive oxygen species.
© 2022 Chinese Pharmaceutical Association and Institute of Materia Medica, Chinese Academy of Medical Sciences. Production and hosting by Elsevier B.V.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- Vezzani A., Balosso S., Ravizza T. Neuroinflammatory pathways as treatment targets and biomarkers in epilepsy. Nat Rev Neurol. 2019;15:459–472. - PubMed
-
- Drion C.M., van Scheppingen J., Arena A., Geijtenbeek K.W., Kooijman L., van Vliet E.A., et al. Effects of rapamycin and curcumin on inflammation and oxidative stress in vitro and in vivo—in search of potential anti-epileptogenic strategies for temporal lobe epilepsy. J Neuroinflammation. 2018;15:212. - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
