

doi: 10.1158/0008-5472.CAN-22-3030.
Extrachromosomal Amplification of Human Papillomavirus Episomes Is a Mechanism of Cervical Carcinogenesis
Affiliations
- PMID: 36971511
- PMCID: PMC10239328
- DOI: 10.1158/0008-5472.CAN-22-3030
Item in Clipboard
Extrachromosomal Amplification of Human Papillomavirus Episomes Is a Mechanism of Cervical Carcinogenesis
Cancer Res.
.
Abstract
Multimers of the HPV genome are generated in cervical tumors replicating as extrachromosomal episomes, which is associated with deletion and rearrangement of the HPV genome and provides a mechanism for oncogenesis without integration.
©2023 American Association for Cancer Research.
Conflict of interest statement
Conflicts of interest
The authors declare no potential conflicts of interest.
Figures
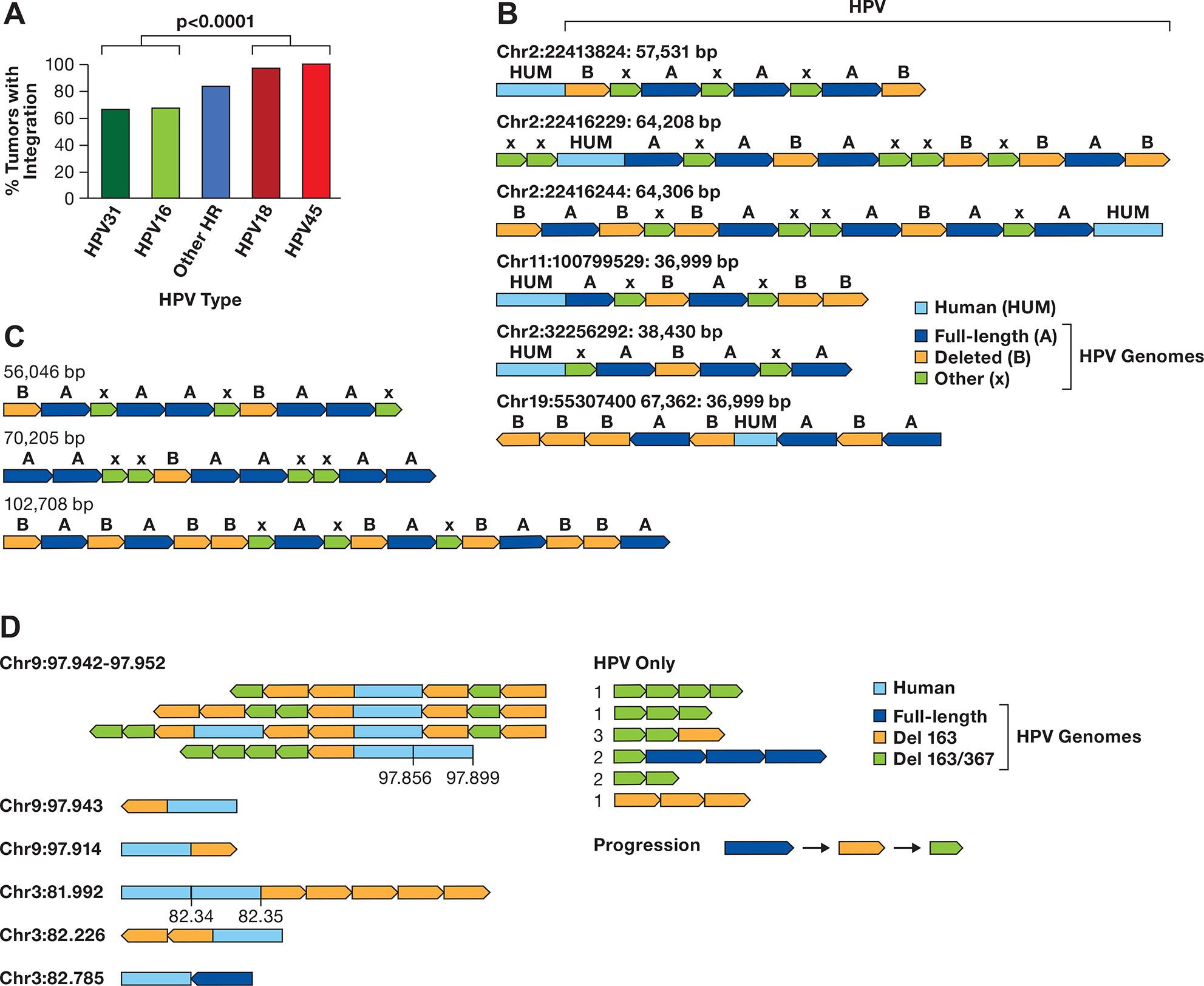
A. The integration frequency of the predominant carcinogenic HPV types is shown using combined data from TCGA and Guatemalan tumors (4,27). The other high-risk (HR) types are combined. The number of samples is HPV31 (N=3), HPV16 (N=170), Other HR (N=43) includes 13 HPV types, HPV18 (N=32), and HPV45 (N=17). The comparision of HPV16/31 to HPV16/45 is significant, P> 0.0001 byt Fishers exact test. B. A diagram of contigs of long DNA sequence reads in CaSki cells aligning to the human genome is shown. Human DNA sequences are in light blue, the full-length HPV16 genome A is dark blue, a 6.5 kb deleted genome derived from genome A is orange, and other smaller HPV16 fragments of varying size are green (X). The arrows show the direction of the HPV genome segments. The location of the human junction and size of the contig is shown. C. HPV only reads from CaSki cells, key as in panel B. D. Contigs of SCC152 HPV16 multimers are shown, with the human/HPV junction location on the left in Mb. For reads with an additional human segment, those coordinates are given above the contig. For HPV-only reads, the number of times that structure was found are shown to the left of the read diagram. A suggested progression in the evolution of the single and double deleted HPV16 genomes is shown.
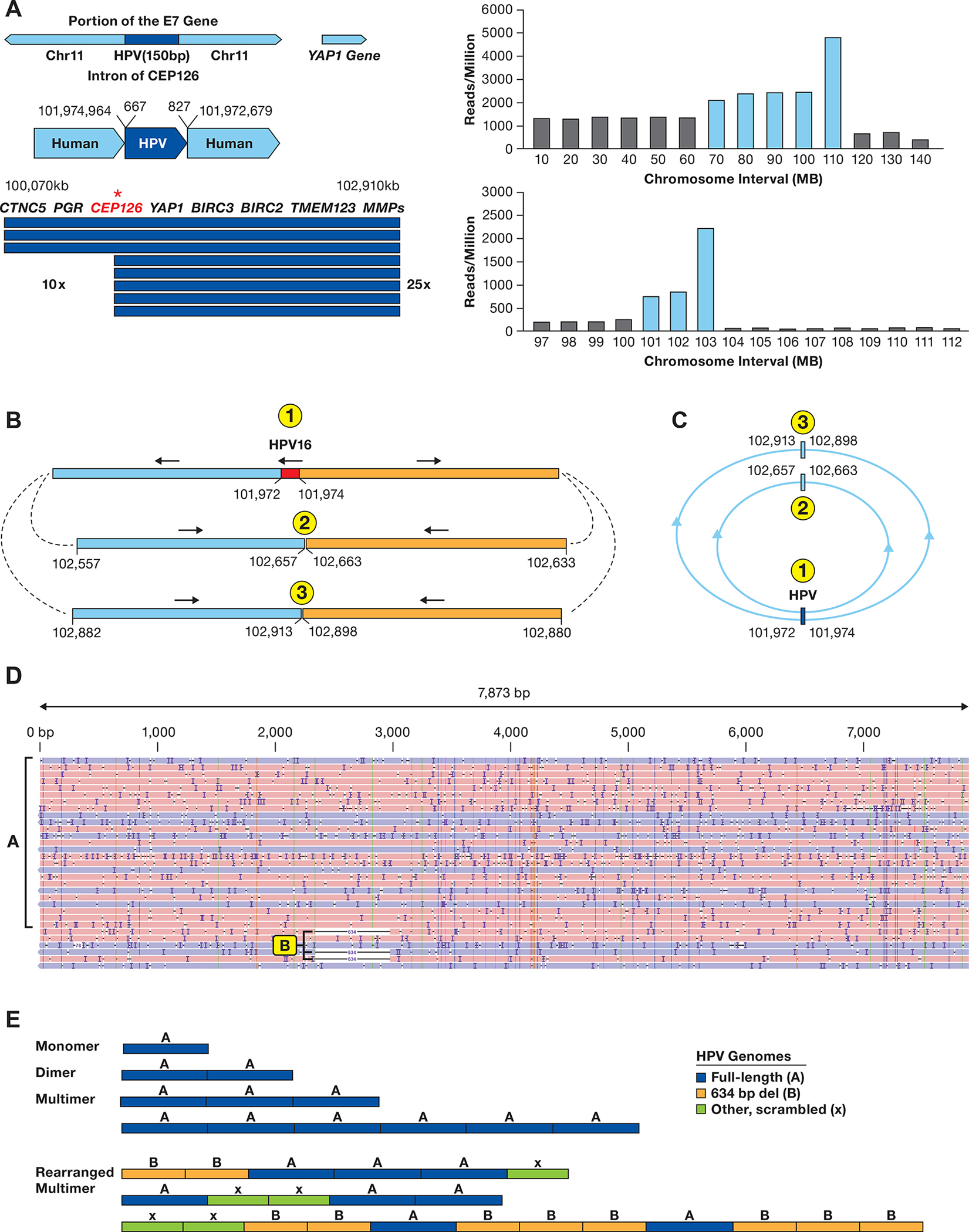
A. The integration locus on chromosome 11 in SNU-1000 is shown along with the fragment of HPV16 from 667–827 bp of the HPV16 genome. The arrow displays the site of integration in the CEP126 gene and the amplified 2.9 Mb region. Below are read counts normalized to reads/million total reads separately in 10 and 1 Mb intervals across chromosome 11 and the 97–112 Mb region. Blue bars represent regions amplified as compared to the rest of the genome. Most other chromosomes do not show such drastic alterations in read depth (Supplementary Figure S9). B. Long-read contigs containing the sequences flanking the integration site are shown along with the direction of the flanking human segments. Also shown are contigs of the two human-human junctions supported by multiple reads joining chr11:102,657 to 102,663 kb and chr11:102,913 to 102,898 kb. C. A model of circular structures that would be consistent with the read junctions, forming 1.4 and 1.9 Mb circles. D. SNU-1000 WGS reads mapped to the HPV16 genome show insertions of multimers of ~7.9 kb, as well as 634 bp deletion genomes and rearranged genomes (arrows). E. Representative SNU-1000 HPV-only reads corresponding to monomer, dimers, and multimer forms of the full-length HPV genome. In addition, multimers containing concatemers of full-length, 634 bp deleted, and other HPV fragments are displayed.
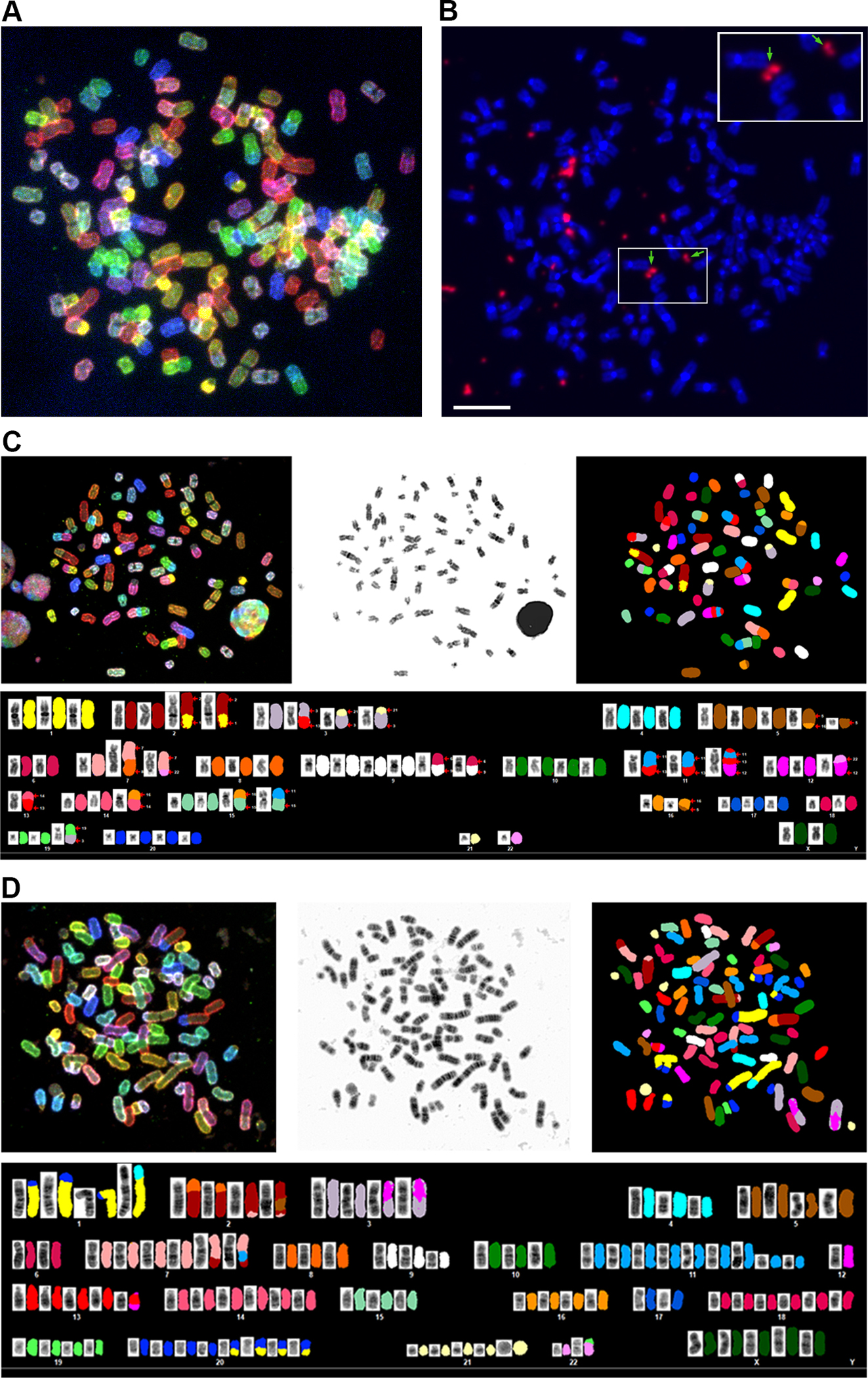
A. A representative spectral karyotype of SNU-1000 cell is shown revealing a near-triploid chromosome content. B. Fluorescent in situ hybridization (FISH) of the same metaphase with a pan-HPV probe reveals intense signals for HPV (in orange) in extrachromosomal regions between or adjacent to chromosomes. Arrows and the inset show HPV signals resembling double minute-like structures. The scale bar is 10 um. C. SKY of SNU-1000 displaying translocation of several chromosomes, including chromosome 11 in the region of the YAP1 amplification. D, SKY of SNU-1245, a near-tetraploid cell line with. Details of metaphases and additional images are in Supplementary Figures S3, S4 and Supplemental Tables 6–8).
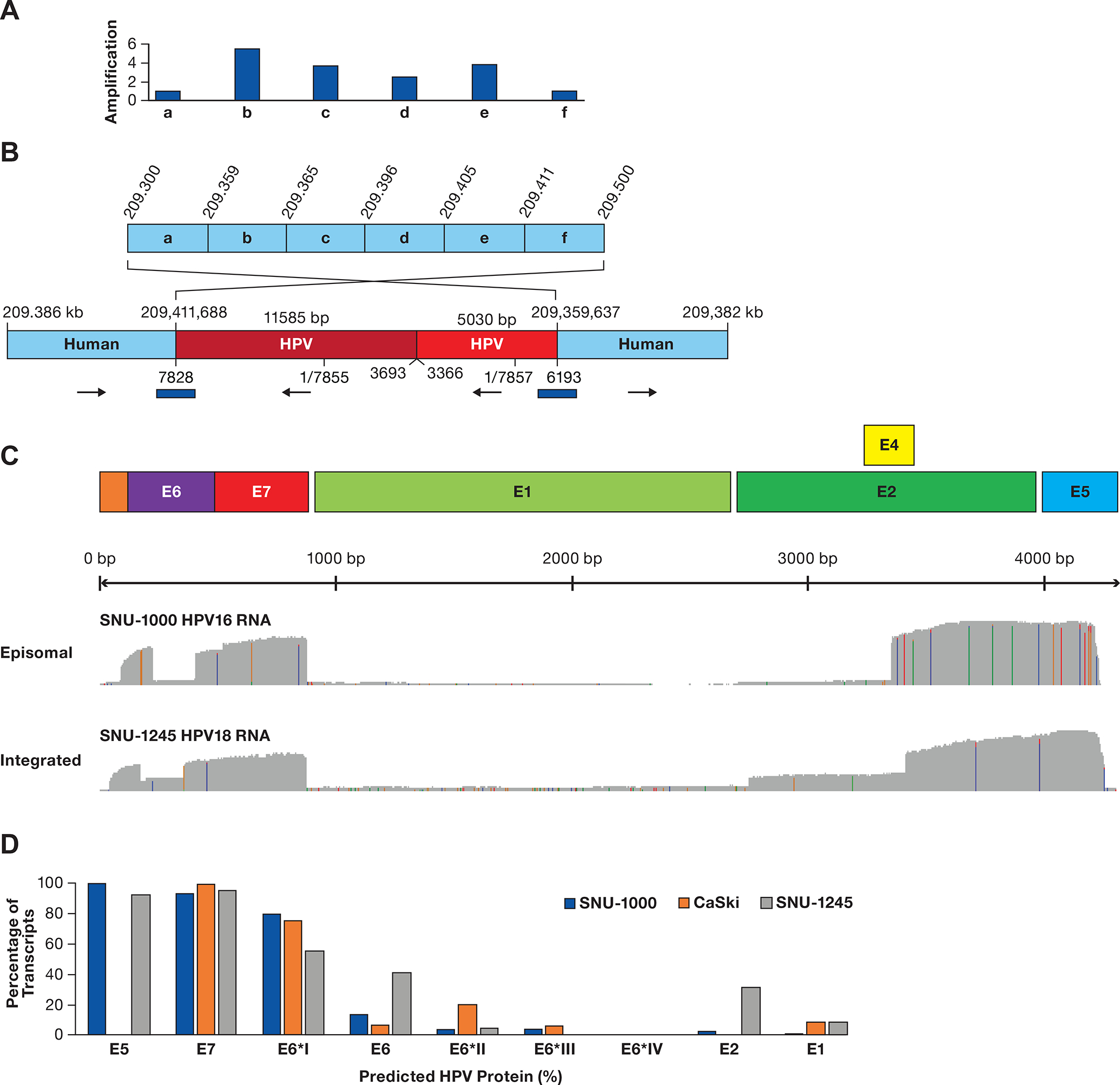
A. A diagram of a region of chromosome 1q24.2 is shown divided into six segments (a-f). Above the diagram are copy number values normalized to segments a and f. B. A contig supported by multiple long reads obtained from ONT WGS, CRISPR targeted sequencing, and adaptive sampling is shown. The two human regions are rearranged, consistent with integration at 209,411,668 bp and looping back to 209,359,637 bp. Human coordinates are above the contig and HPV18 coordinates are below. The total HPV contig is 16.6 kb. The blue bars represent junctions confirmed by PCR and Sanger sequencing. C. Plots of full-length direct cDNA sequencing are shown, displaying an abundant expression of the E6/E7 gene regions with frequent splicing of E6 and deficient expression of the E1/E2 gene region. D. The estimated level of HPV proteins is shown based on content and abundance of transcripts in Supplementary Table S9 and Supplementary Figure S5, assuming transcripts produce equal amounts of protein.
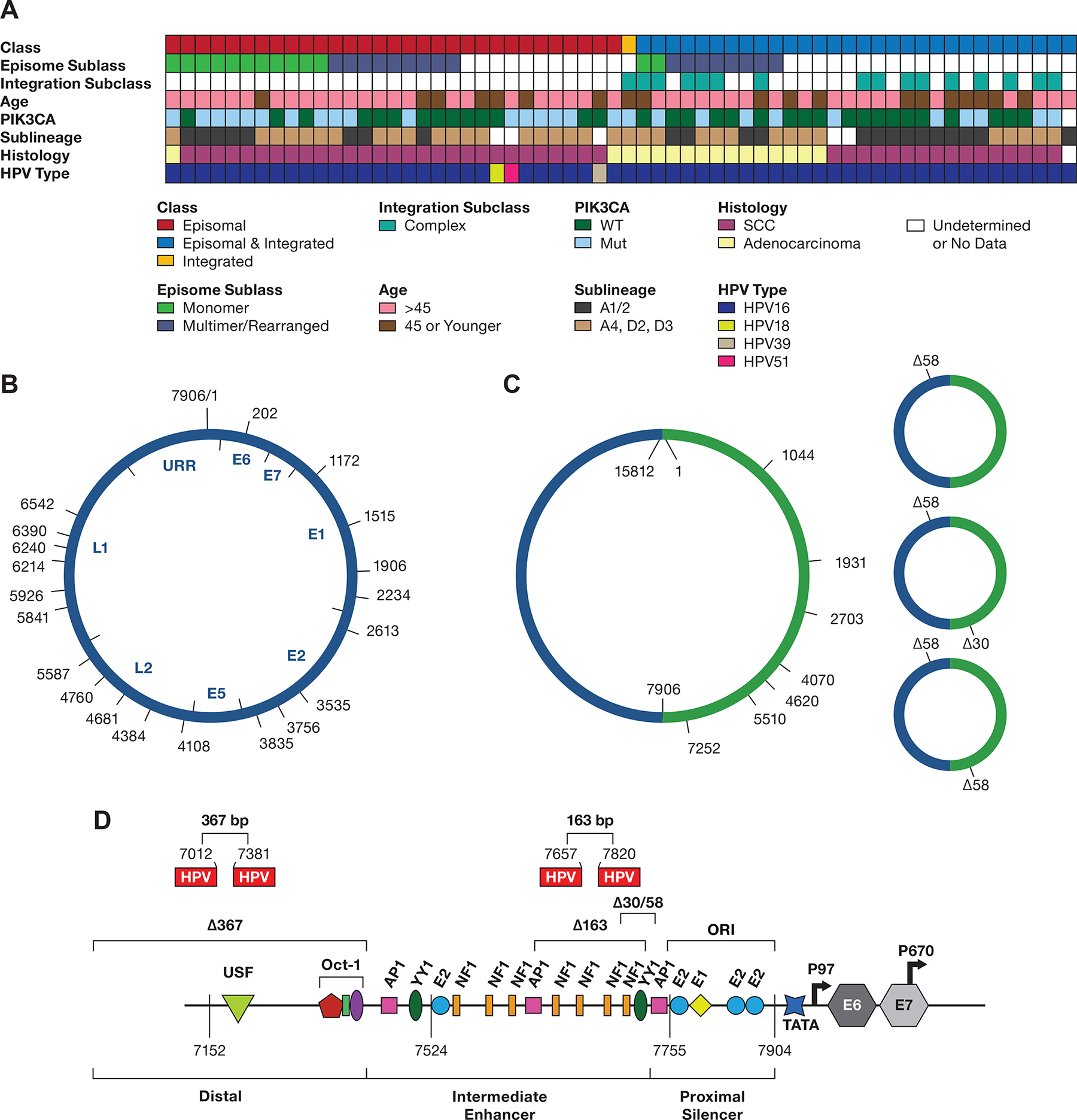
A. The integration class and subclass, episomal subclass, age range, PIK3CA mutation status, HPV type and HPV16 sublineage, and histology are shown. SCC, squamous cell carcinoma; WT, wild type; Int, integration. Blanks represent samples not able to be classified. B. Diagram of the 7906 bp HPV16 genome DNA, displaying the start site of DNA sequence reads from 13 tumors containing a complete or nearly complete monomer genome sequence. The position of the HPV16 genes and URR are shown inside the diagram. C, A diagram of HPV16 dimers from three tumors showing the start position of reads with complete or nearly complete copies of two tandem HPV16 genomes (Supplemental Table 10). D, the location of 30 and 58 bp deletions in the URR occurring in dimer reads from tumor T393.The location of the 163 and 367 bp deletions in the SCC152 cell line is shown, as well as their locations in the HPV16 upstream regulatory region. The 367 bp deletion removes most of the distal region, and the 163 bp deletion removes four NF1 binding sites and two YY1 binding sites (only one is shown) in the Intermediate Enhancer region. Shown are the binding sites for the viral E1 and E2 proteins and the transcription factors OCT1, AP1, YY1, and NF1. ORI, the origin of replication; TATA, TATA-binding site; P97, major promoter; P670, minor promoter. The locations of the coding region of the E6 and E7 genes are indicated
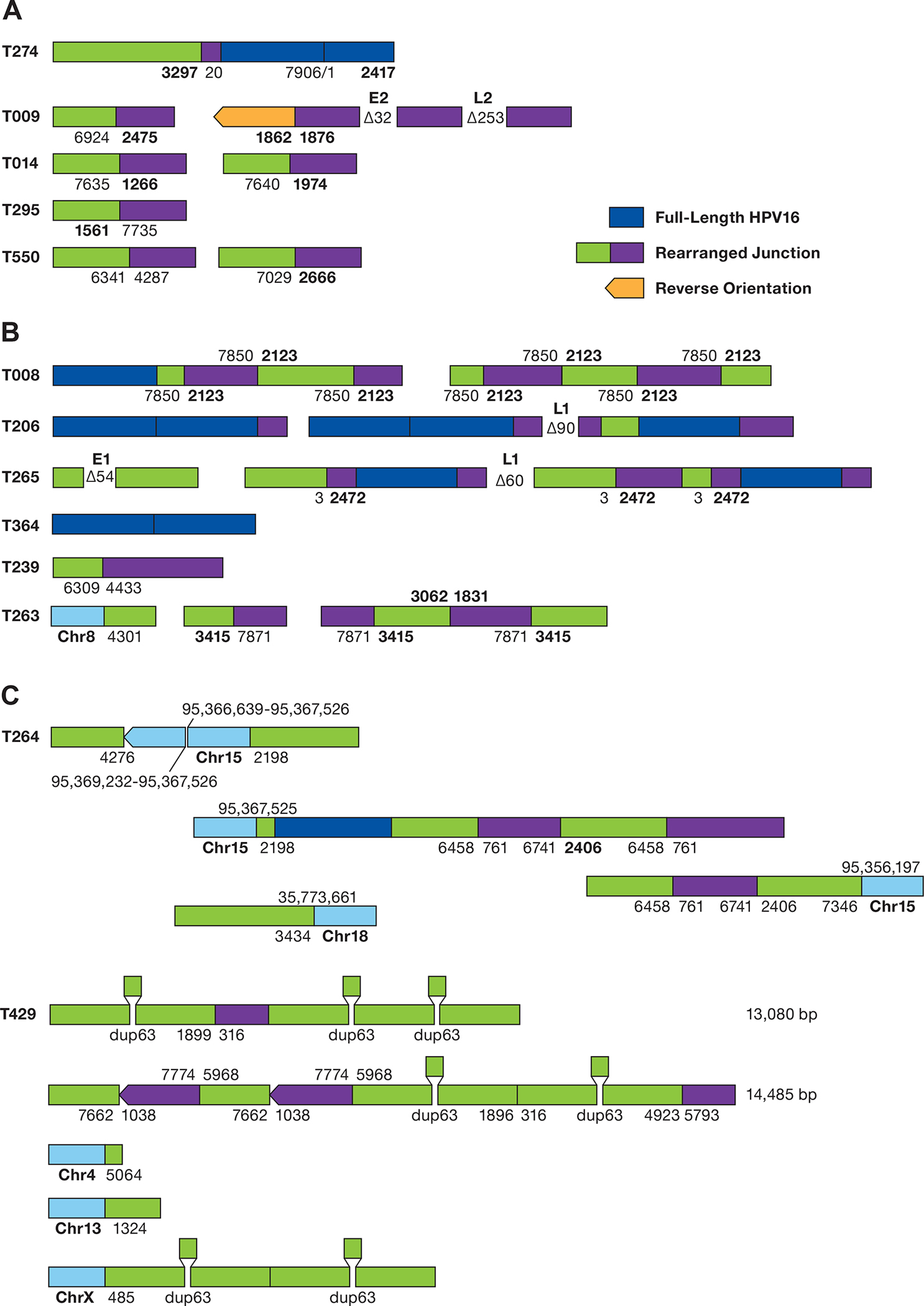
A. The structure of rearranged HPV-only reads in EP tumors is shown. The numbers represent the position of the HPV16 genome at the site of the breakpoint. Deletions are displayed between segments. Unless shown, all HPV sequences are arranged in the same orientation. B. Structure of HPV reads in putative type 2 tumors. Regions of human DNA are shown in light blue. C. The structure of HPV reads in tumor T429 is shown. HPV16 sequence breakpoints are shown joining chromosomes 4, 13, and X to HPV sequences. The position of a 63 bp duplication is shown. The top 2 diagrams represent independent sequence reads of 13,085 and 14,485 bp.
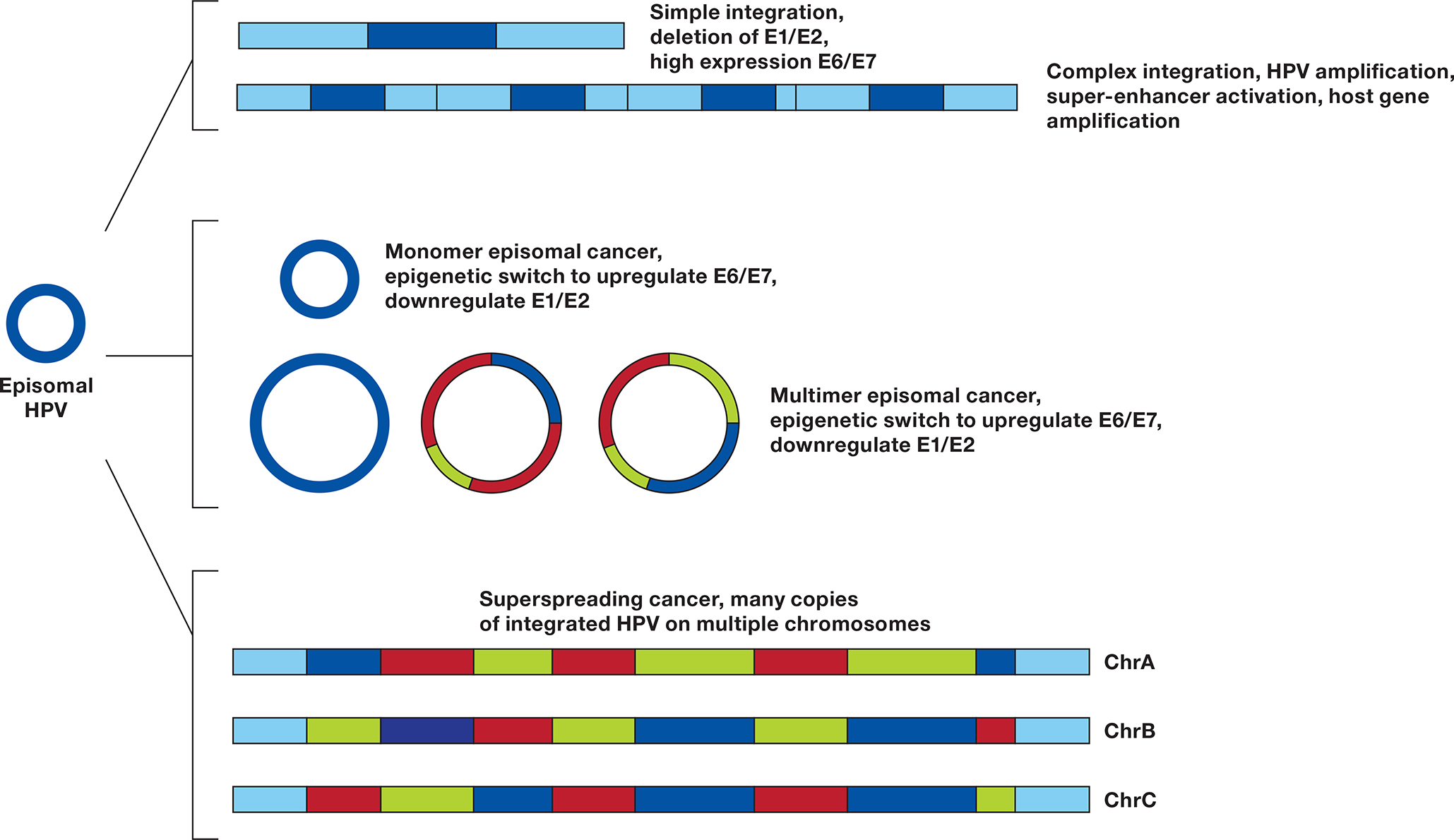
An HPV episome can undergo a simple integration, deleting the E1 and E2 genes, inhibiting DNA synthesis at the HPV origin of replication (ORI) and upregulating E6 and E7. If during integration E1 and E2 are not suppressed, a local amplification can case a complex integration leading to multiple copies of HPV and flanking DNA, resulting in higher expression of E6 and E7, formation of a super-enhancer activating flanking genes (23), and potentially amplifying a carcinogenic host gene. Monomer episomal cancer occurs when the genome remains an episome but undergoes an epigenetic switch to upregulate E6/E7. Aberrant episome replication can lead to Multimer episomal cancer in which deletions and rearrangements in the E1/E2 genes or URR lead to high E6/E7 and low E1/E2 expression. Insertion of multimer episomes into multiple genomic locations give rise to HPV superspreading cancer.
References
-
- Schiffman M, Doorbar J, Wentzensen N, de Sanjose S, Fakhry C, Monk BJ, et al. Carcinogenic human papillomavirus infection. Nat Rev Dis Primers 2016;2:16086. - PubMed
-
- Walboomers JM, Jacobs MV, Manos MM, Bosch FX, Kummer JA, Shah KV, et al. Human papillomavirus is a necessary cause of invasive cervical cancer worldwide. J Pathol 1999;189:12–9 - PubMed
-
- Schiffman M, Castle PE. The promise of global cervical-cancer prevention. N Engl J Med 2005;353:2101–4 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
