

An associative memory Hamiltonian model for DNA and nucleosomes
- PMID: 36972316
- PMCID: PMC10079229
- DOI: 10.1371/journal.pcbi.1011013
An associative memory Hamiltonian model for DNA and nucleosomes
Abstract
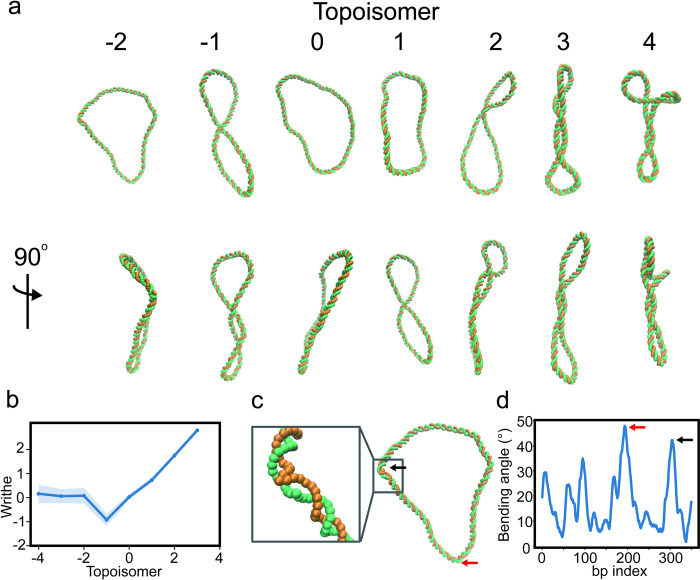
A model for DNA and nucleosomes is introduced with the goal of studying chromosomes from a single base level all the way to higher-order chromatin structures. This model, dubbed the Widely Editable Chromatin Model (WEChroM), reproduces the complex mechanics of the double helix including its bending persistence length and twisting persistence length, and the temperature dependence of the former. The WEChroM Hamiltonian is composed of chain connectivity, steric interactions, and associative memory terms representing all remaining interactions leading to the structure, dynamics, and mechanical characteristics of the B-DNA. Several applications of this model are discussed to demonstrate its applicability. WEChroM is used to investigate the behavior of circular DNA in the presence of positive and negative supercoiling. We show that it recapitulates the formation of plectonemes and of structural defects that relax mechanical stress. The model spontaneously manifests an asymmetric behavior with respect to positive or negative supercoiling, similar to what was previously observed in experiments. Additionally, we show that the associative memory Hamiltonian is also capable of reproducing the free energy of partial DNA unwrapping from nucleosomes. WEChroM is designed to emulate the continuously variable mechanical properties of the 10nm fiber and, by virtue of its simplicity, is ready to be scaled up to molecular systems large enough to investigate the structural ensembles of genes. WEChroM is implemented in the OpenMM simulation toolkits and is freely available for public use.
Copyright: © 2023 Lu et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



References
-
- Wilkins MHF, Stokes AR, Wilson HR. Molecular Structure of Nucleic Acids: Molecular Structure of Deoxypentose Nucleic Acids. Nature. 1953. Apr;171(4356):738–40. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
