

Linking prokaryotic genome size variation to metabolic potential and environment
- PMID: 36973336
- PMCID: PMC10042847
- DOI: 10.1038/s43705-023-00231-x
Linking prokaryotic genome size variation to metabolic potential and environment
Abstract
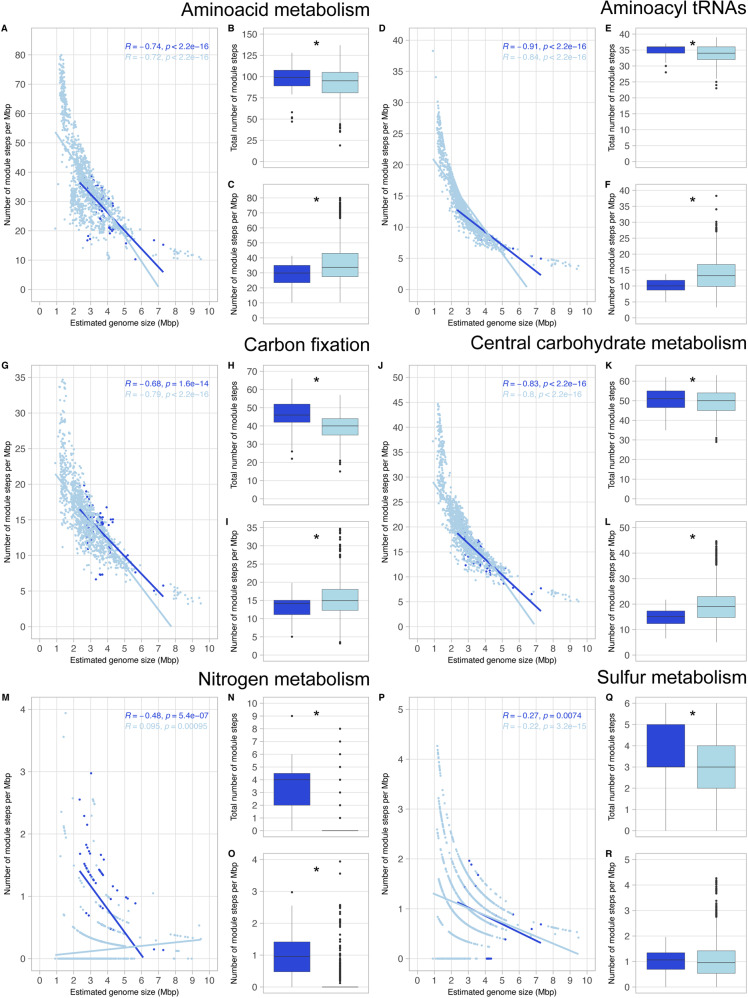
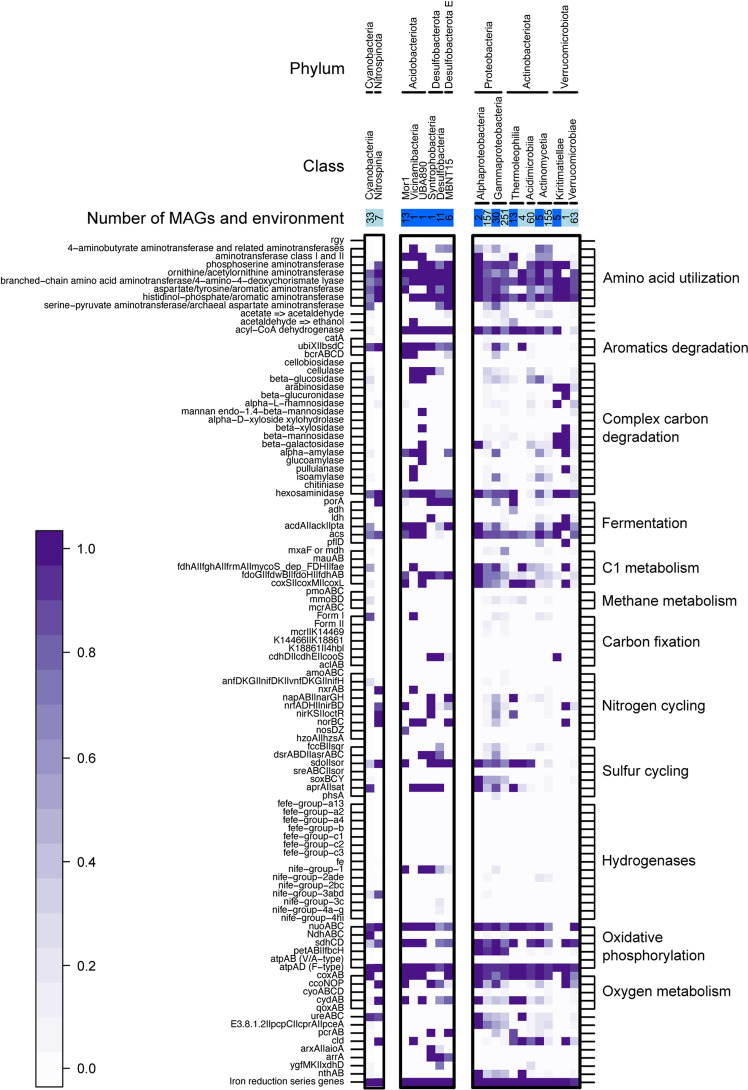
While theories and models have appeared to explain genome size as a result of evolutionary processes, little work has shown that genome sizes carry ecological signatures. Our work delves into the ecological implications of microbial genome size variation in benthic and pelagic habitats across environmental gradients of the brackish Baltic Sea. While depth is significantly associated with genome size in benthic and pelagic brackish metagenomes, salinity is only correlated to genome size in benthic metagenomes. Overall, we confirm that prokaryotic genome sizes in Baltic sediments (3.47 Mbp) are significantly bigger than in the water column (2.96 Mbp). While benthic genomes have a higher number of functions than pelagic genomes, the smallest genomes coded for a higher number of module steps per Mbp for most of the functions irrespective of their environment. Some examples of this functions are amino acid metabolism and central carbohydrate metabolism. However, we observed that nitrogen metabolism was almost absent in pelagic genomes and was mostly present in benthic genomes. Finally, we also show that Bacteria inhabiting Baltic sediments and water column not only differ in taxonomy, but also in their metabolic potential, such as the Wood-Ljungdahl pathway or the presence of different hydrogenases. Our work shows how microbial genome size is linked to abiotic factors in the environment, metabolic potential and taxonomic identity of Bacteria and Archaea within aquatic ecosystems.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
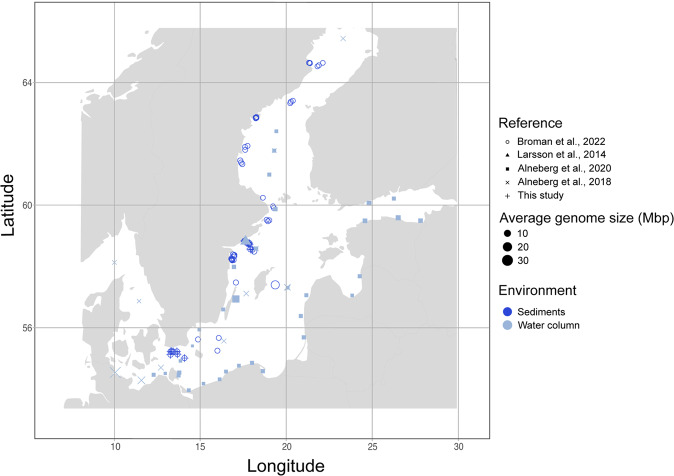
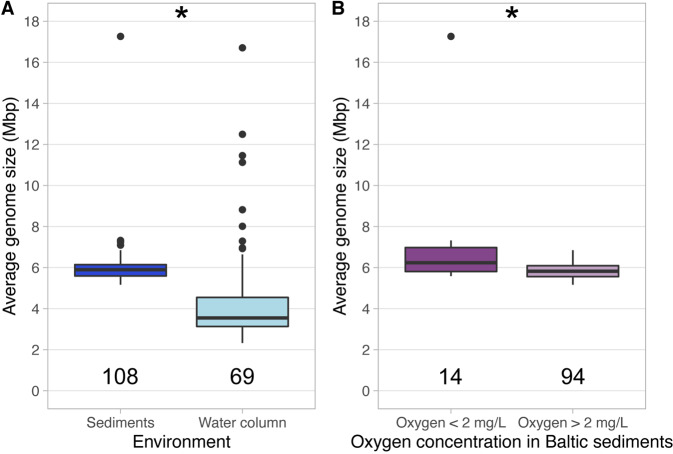
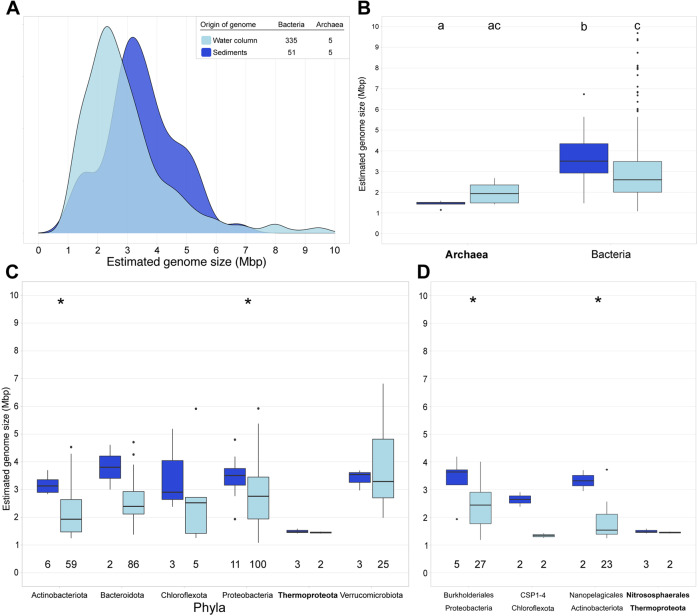
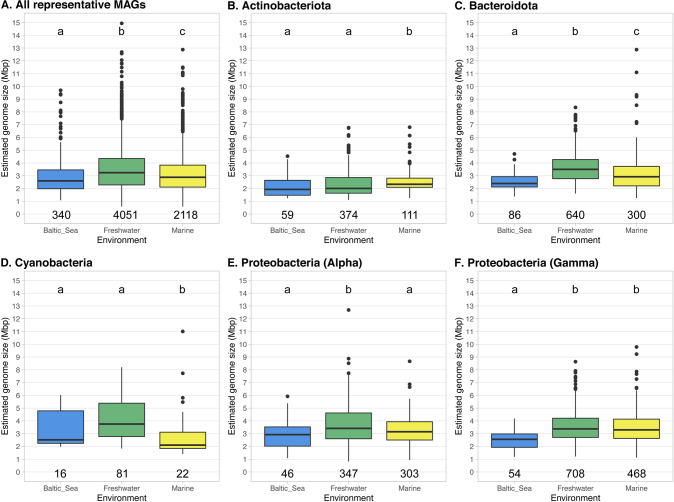
References
-
- Rodríguez-Gijón A, Nuy JK, Mehrshad M, Buck M, Schulz F, Woyke T, et al. A genomic perspective across Earth’s microbiomes reveals that genome size in Archaea and Bacteria is linked to ecosystem type and trophic strategy. Front Microbiol. 2022;12:761869. doi: 10.3389/fmicb.2021.761869. - DOI - PMC - PubMed
-
- Lynch M. Streamlining and simplification of microbial genome architecture. Annu Rev Microbiol. 2006;60:327–49. - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
