

The NLR gene family: from discovery to present day
- PMID: 36973360
- PMCID: PMC11171412
- DOI: 10.1038/s41577-023-00849-x
The NLR gene family: from discovery to present day
Erratum in
-
Author Correction: The NLR gene family: from discovery to present day.Nat Rev Immunol. 2023 Jul;23(7):472. doi: 10.1038/s41577-023-00891-9. Nat Rev Immunol. 2023. PMID: 37198352 No abstract available.
Abstract
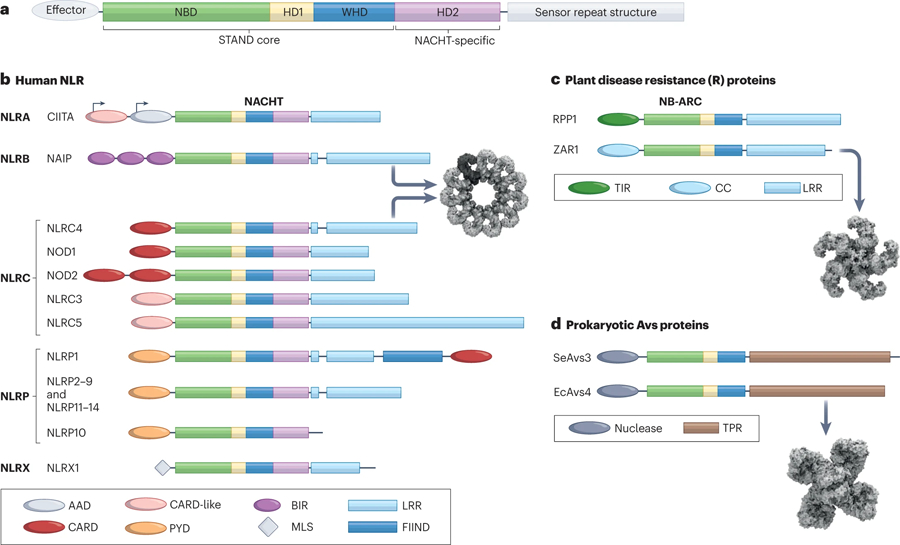
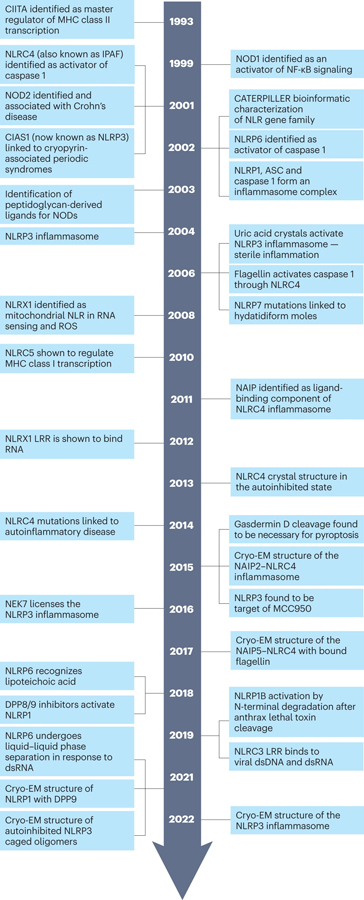
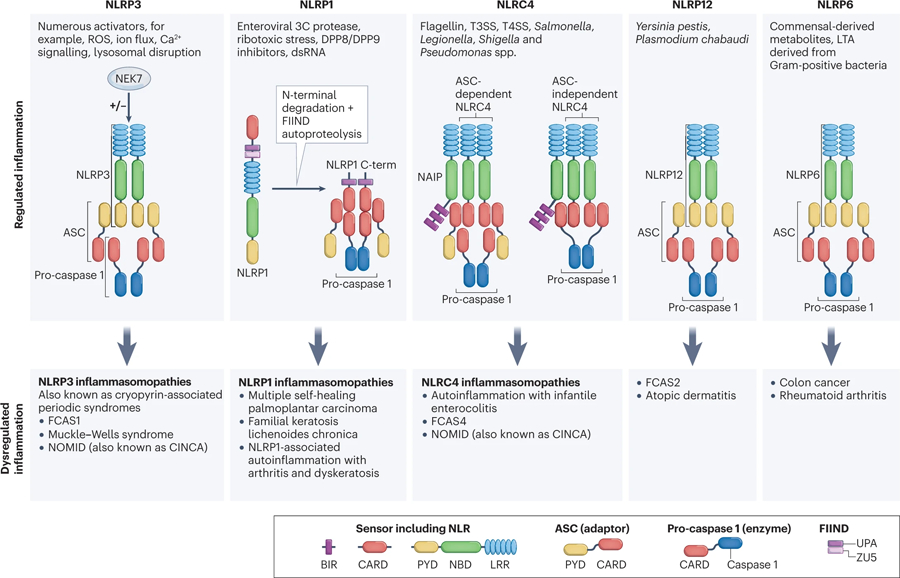
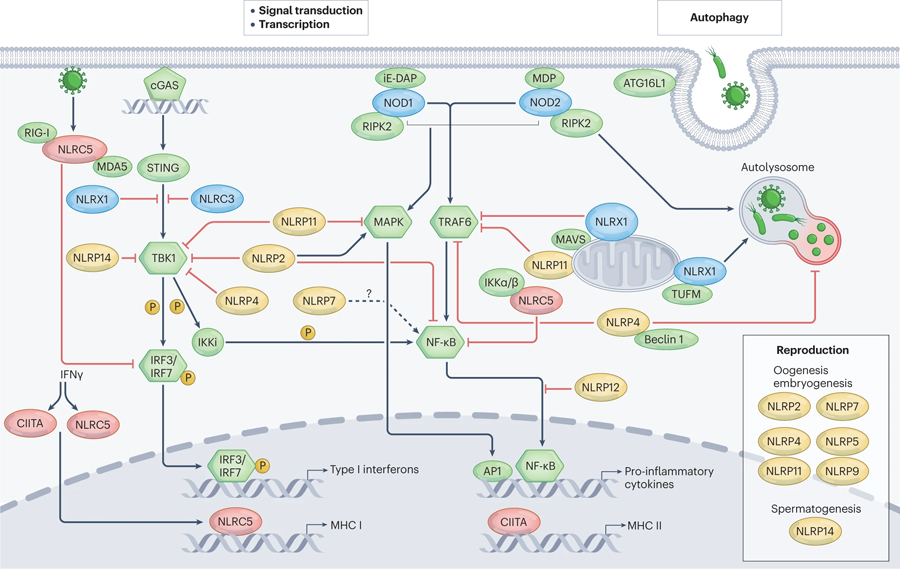
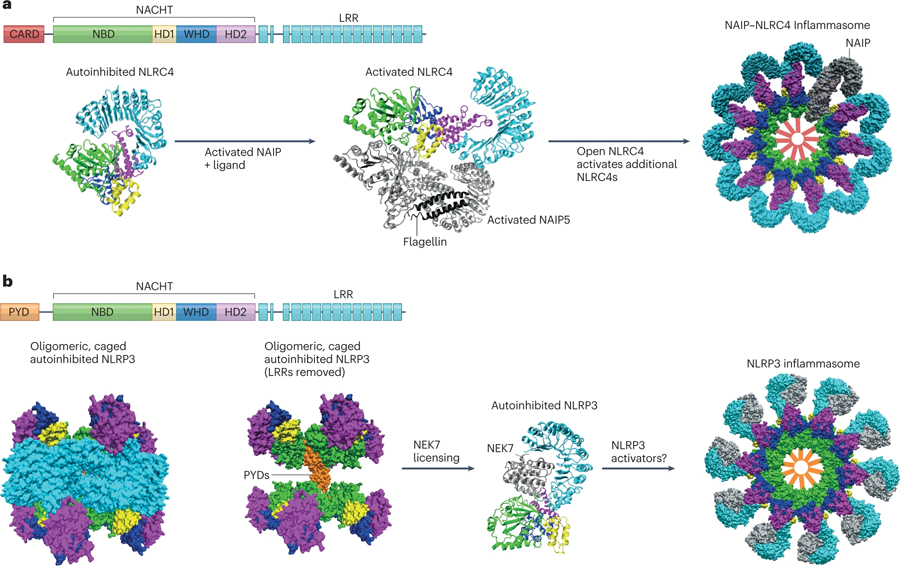
The mammalian NLR gene family was first reported over 20 years ago, although several genes that were later grouped into the family were already known at that time. Although it is widely known that NLRs include inflammasome receptors and/or sensors that promote the maturation of caspase 1, IL-1β, IL-18 and gasdermin D to drive inflammation and cell death, the other functions of NLR family members are less well appreciated by the scientific community. Examples include MHC class II transactivator (CIITA), a master transcriptional activator of MHC class II genes, which was the first mammalian NBD-LRR-containing protein to be identified, and NLRC5, which regulates the expression of MHC class I genes. Other NLRs govern key inflammatory signalling pathways or interferon responses, and several NLR family members serve as negative regulators of innate immune responses. Multiple NLRs regulate the balance of cell death, cell survival, autophagy, mitophagy and even cellular metabolism. Perhaps the least discussed group of NLRs are those with functions in the mammalian reproductive system. The focus of this Review is to provide a synopsis of the NLR family, including both the intensively studied and the underappreciated members. We focus on the function, structure and disease relevance of NLRs and highlight issues that have received less attention in the NLR field. We hope this may serve as an impetus for future research on the conventional and non-conventional roles of NLRs within and beyond the immune system.
© 2023. Springer Nature Limited.
Conflict of interest statement
Competing Interests
J. Ting is a co-founder of IMMvention Therapeutix.
Figures
References
-
- Steimle V, Otten LA, Zufferey M & Mach B Complementation cloning of an MHC class II transactivator mutated in hereditary MHC class II deficiency (or bare lymphocyte syndrome). Cell 75, 135–146 (1993). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
