

Common seed dispersers contribute most to the persistence of a fleshy-fruited tree
- PMID: 36973362
- PMCID: PMC10043030
- DOI: 10.1038/s42003-023-04647-y
Common seed dispersers contribute most to the persistence of a fleshy-fruited tree
Abstract
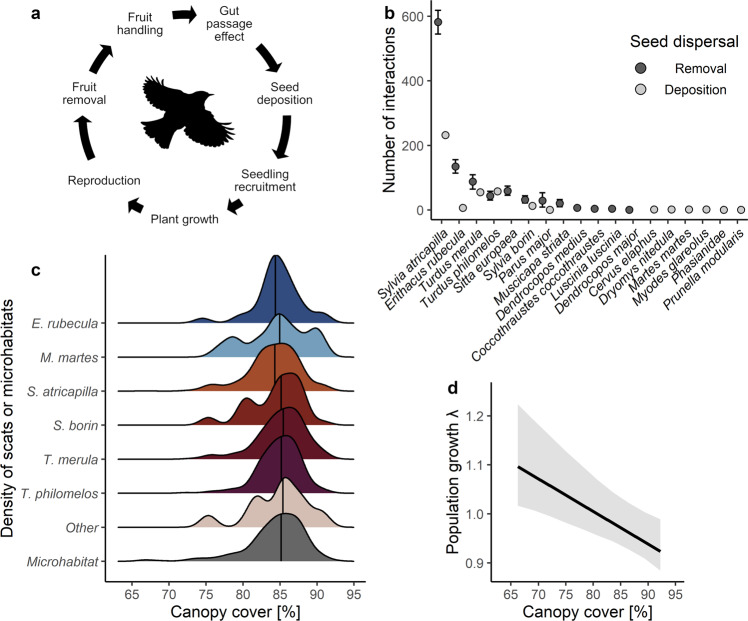
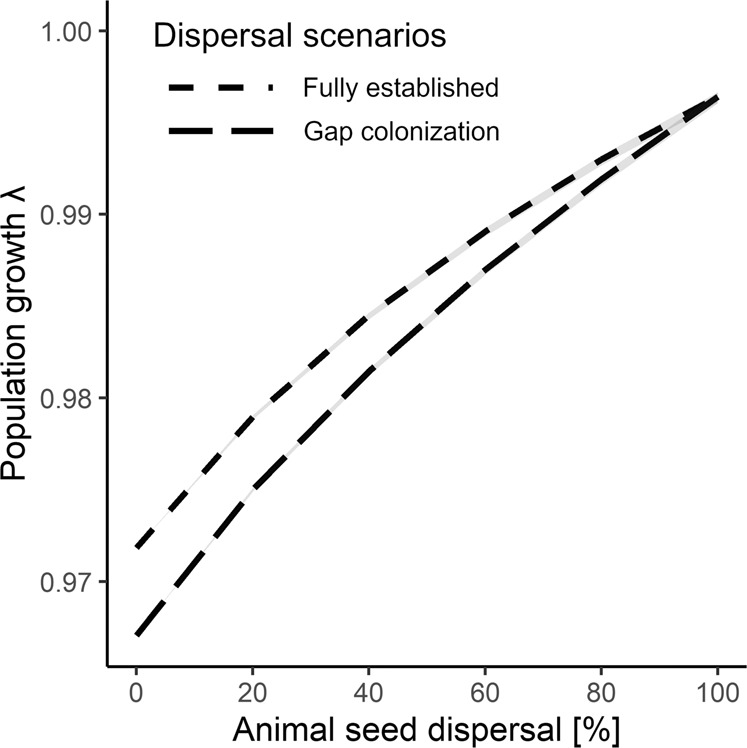
Mutualistic interactions are by definition beneficial for each contributing partner. However, it is insufficiently understood how mutualistic interactions influence partners throughout their lives. Here, we used animal species-explicit, microhabitat-structured integral projection models to quantify the effect of seed dispersal by 20 animal species on the full life cycle of the tree Frangula alnus in Białowieża Forest, Eastern Poland. Our analysis showed that animal seed dispersal increased population growth by 2.5%. The effectiveness of animals as seed dispersers was strongly related to the interaction frequency but not the quality of seed dispersal. Consequently, the projected population decline due to simulated species extinction was driven by the loss of common rather than rare mutualist species. Our results support the notion that frequently interacting mutualists contribute most to the persistence of the populations of their partners, underscoring the role of common species for ecosystem functioning and nature conservation.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Harvey E, Gounand I, Ward CL, Altermatt F. Bridging ecology and conservation: from ecological networks to ecosystem function. J. Appl. Ecol. 2017;54:371–379. doi: 10.1111/1365-2664.12769. - DOI
-
- Vázquez DP, Morris WF, Jordano P. Interaction frequency as a surrogate for the total effect of animal mutualists on plants. Ecol. Lett. 2005;8:1088–1094. doi: 10.1111/j.1461-0248.2005.00810.x. - DOI
-
- Winfree, R. in Unsolved Problems in Ecology 338–354 (Princeton University Press, 2020).
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
