

Metabolomic Analysis Demonstrates the Impacts of Polyketide Synthases PKS14 and PKS15 on the Production of Beauvericins, Bassianolide, Enniatin A, and Ferricrocin in Entomopathogen Beauveria bassiana
- PMID: 36984865
- PMCID: PMC10057652
- DOI: 10.3390/metabo13030425
Metabolomic Analysis Demonstrates the Impacts of Polyketide Synthases PKS14 and PKS15 on the Production of Beauvericins, Bassianolide, Enniatin A, and Ferricrocin in Entomopathogen Beauveria bassiana
Abstract
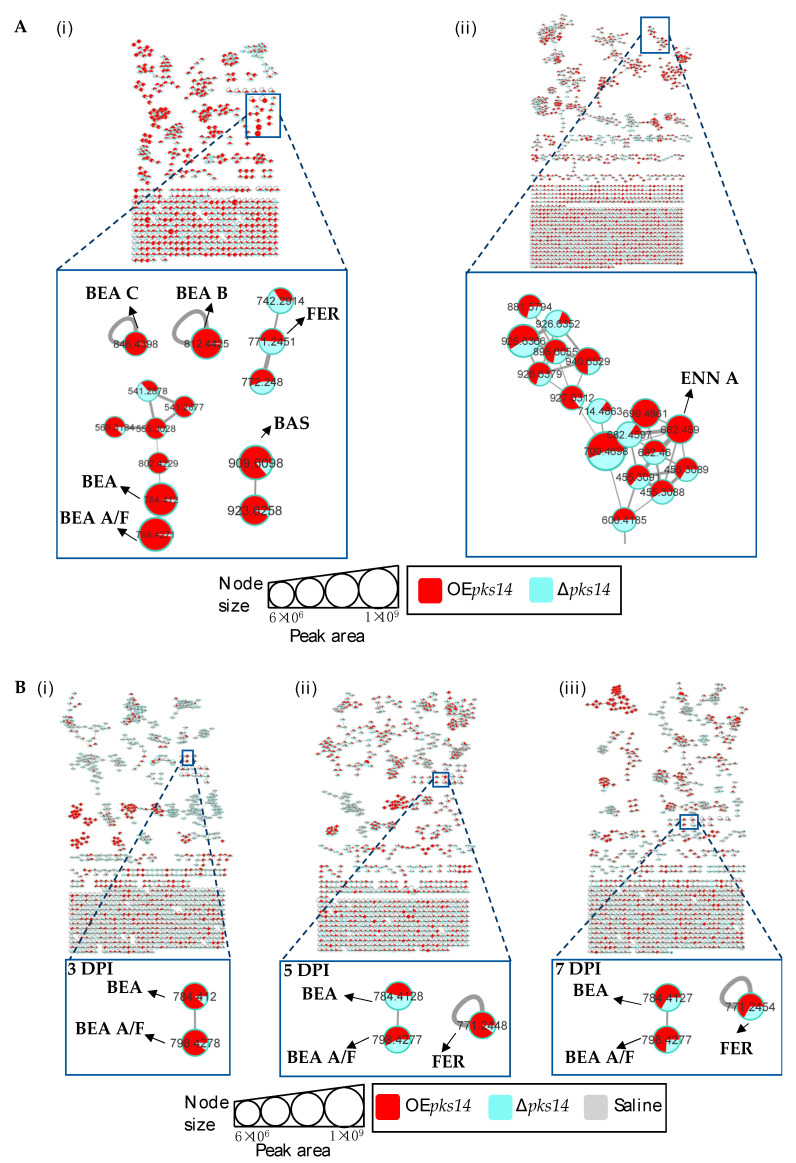
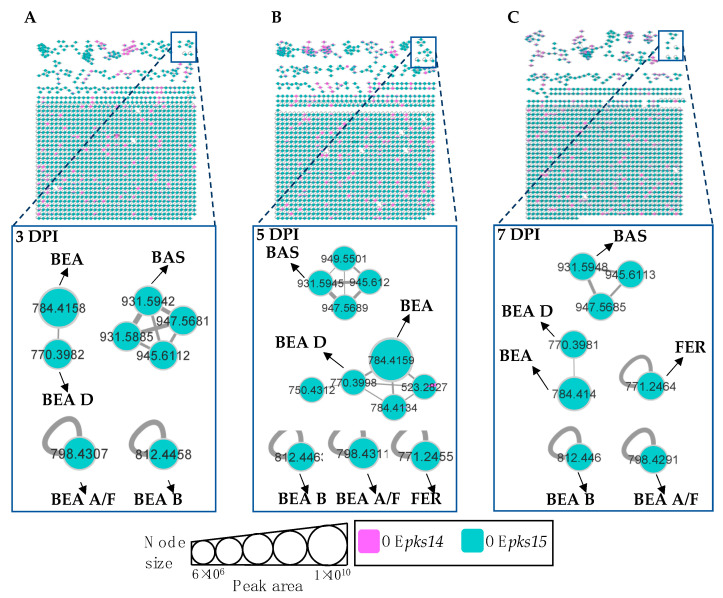
Beauveria bassiana is a globally distributed entomopathogenic fungus that produces various secondary metabolites to support its pathogenesis in insects. Two polyketide synthase genes, pks14 and pks15, are highly conserved in entomopathogenic fungi and are important for insect virulence. However, understanding of their mechanisms in insect pathogenicity is still limited. Here, we overexpressed these two genes in B. bassiana and compared the metabolite profiles of pks14 and pks15 overexpression strains to those of their respective knockout strains in culture and in vivo using tandem liquid chromatography-mass spectrometry (LC-MS/MS) with Global Natural Products Social Molecular Networking (GNPS). The pks14 and pks15 clusters exhibited crosstalk with biosynthetic clusters encoding insect-virulent metabolites, including beauvericins, bassianolide, enniatin A, and the intracellular siderophore ferricrocin under certain conditions. These secondary metabolites were upregulated in the pks14-overexpressing strain in culture and the pks15-overexpressing strain in vivo. These data suggest that pks14 and pks15, their proteins or their cluster components might be directly or indirectly associated with key pathways in insect pathogenesis of B. bassiana, particularly those related to secondary metabolism. Information about interactions between the polyketide clusters and other biosynthetic clusters improves scientific understanding about crosstalk among biosynthetic pathways and mechanisms of pathogenesis.
Keywords: Beauveria bassiana; GNPS; molecular networking; nonribosomal peptides; polyketide synthase.
Conflict of interest statement
There are no conflicts of interest to declare.
Figures





Similar articles
-
Transcriptomic insights into the interplay between polyketide biosynthesis and other secondary metabolite biosynthetic clusters and biological pathways in entomopathogen Beauveria bassiana.Front Microbiol. 2025 May 23;16:1583637. doi: 10.3389/fmicb.2025.1583637. eCollection 2025. Front Microbiol. 2025. PMID: 40485827 Free PMC article.
-
The reducing clade IIb polyketide synthase PKS14 acts as a virulence determinant of the entomopathogenic fungus Beauveria bassiana.FEMS Microbiol Lett. 2018 Aug 1;365(15). doi: 10.1093/femsle/fny131. FEMS Microbiol Lett. 2018. PMID: 29790944
-
Targeted disruption of the polyketide synthase gene pks15 affects virulence against insects and phagocytic survival in the fungus Beauveria bassiana.Fungal Biol. 2017 Aug;121(8):664-675. doi: 10.1016/j.funbio.2017.04.007. Epub 2017 May 6. Fungal Biol. 2017. PMID: 28705395
-
The Entomopathogenic Fungus Beauveria bassiana Shows Its Toxic Side within Insects: Expression of Genes Encoding Secondary Metabolites during Pathogenesis.J Fungi (Basel). 2022 May 7;8(5):488. doi: 10.3390/jof8050488. J Fungi (Basel). 2022. PMID: 35628744 Free PMC article. Review.
-
The Toxins of Beauveria bassiana and the Strategies to Improve Their Virulence to Insects.Front Microbiol. 2021 Aug 26;12:705343. doi: 10.3389/fmicb.2021.705343. eCollection 2021. Front Microbiol. 2021. PMID: 34512581 Free PMC article. Review.
Cited by
-
Isolation and characterization of a native strain of the entomopathogenic fungus Beauveria bassiana for the control of the palm weevil Dynamis borassi (Coleoptera: Curculionidae) in the neotropics.World J Microbiol Biotechnol. 2024 Jul 5;40(9):260. doi: 10.1007/s11274-024-04044-5. World J Microbiol Biotechnol. 2024. PMID: 38967730 Free PMC article.
-
Cross-talk between immunity and behavior: insights from entomopathogenic fungi and their insect hosts.FEMS Microbiol Rev. 2024 Jan 12;48(1):fuae003. doi: 10.1093/femsre/fuae003. FEMS Microbiol Rev. 2024. PMID: 38341280 Free PMC article. Review.
-
Role of microbial pesticides in pest control for lepidopteran: research status and prospect.Biotechnol Lett. 2025 Jun 30;47(4):71. doi: 10.1007/s10529-025-03613-4. Biotechnol Lett. 2025. PMID: 40588701 Review.
-
Transcriptomic insights into the interplay between polyketide biosynthesis and other secondary metabolite biosynthetic clusters and biological pathways in entomopathogen Beauveria bassiana.Front Microbiol. 2025 May 23;16:1583637. doi: 10.3389/fmicb.2025.1583637. eCollection 2025. Front Microbiol. 2025. PMID: 40485827 Free PMC article.
References
-
- Jirakkakul J., Cheevadhanarak S., Punya J., Chutrakul C., Senachak J., Buajarern T., Tanticharoen M., Amnuaykanjanasin A. Tenellin acts as an iron chelator to prevent iron-generated reactive oxygen species toxicity in the entomopathogenic fungus Beauveria bassiana. FEMS Microbiol. Lett. 2015;362:1–8. doi: 10.1093/femsle/fnu032. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
