

Lentiviral Vectors as a Vaccine Platform against Infectious Diseases
- PMID: 36986707
- PMCID: PMC10053212
- DOI: 10.3390/pharmaceutics15030846
Lentiviral Vectors as a Vaccine Platform against Infectious Diseases
Abstract
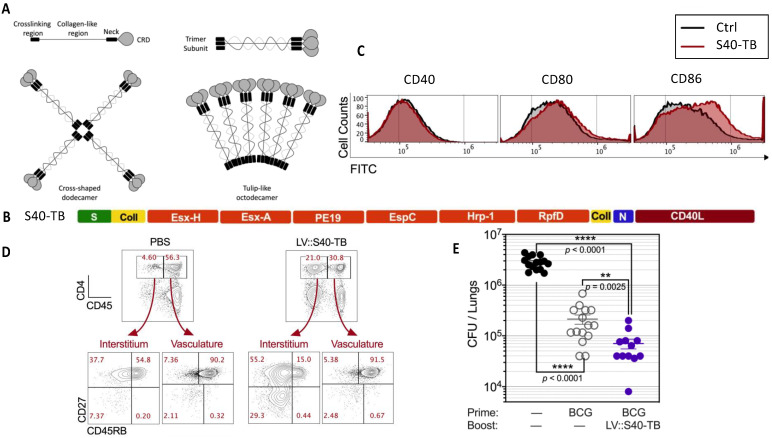
Lentiviral vectors are among the most effective viral vectors for vaccination. In clear contrast to the reference adenoviral vectors, lentiviral vectors have a high potential for transducing dendritic cells in vivo. Within these cells, which are the most efficient at activating naive T cells, lentiviral vectors induce endogenous expression of transgenic antigens that directly access antigen presentation pathways without the need for external antigen capture or cross-presentation. Lentiviral vectors induce strong, robust, and long-lasting humoral, CD8+ T-cell immunity and effective protection against several infectious diseases. There is no pre-existing immunity to lentiviral vectors in the human population and the very low pro-inflammatory properties of these vectors pave the way for their use in mucosal vaccination. In this review, we have mainly summarized the immunological aspects of lentiviral vectors, their recent optimization to induce CD4+ T cells, and our recent data on lentiviral vector-based vaccination in preclinical models, including prophylaxis against flaviviruses, SARS-CoV-2, and Mycobacterium tuberculosis.
Keywords: T-cell vaccines; antigen presentation; inflammation; lentiviral vaccine vectors; major histocompatibility complex; pre-existing anti-vector immunity; tropism for dendritic cells.
Conflict of interest statement
P.C. is the founder and CSO of TheraVectys. LM is a consultant for TheraVectys. KN and FA are employees of TheraVectys. MB and YW declare no competing interest.
Figures
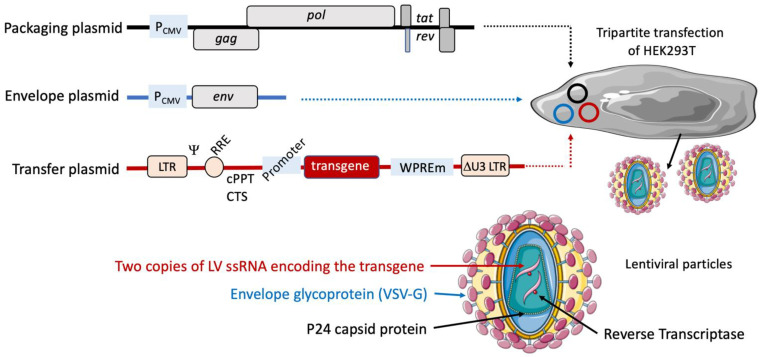
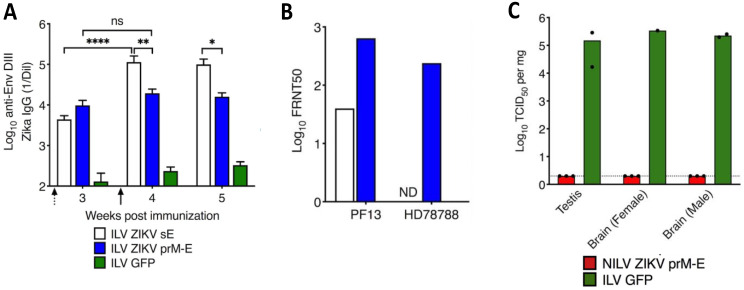
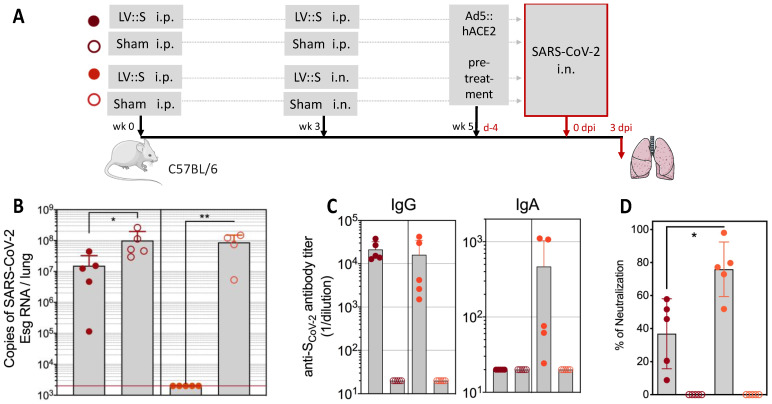
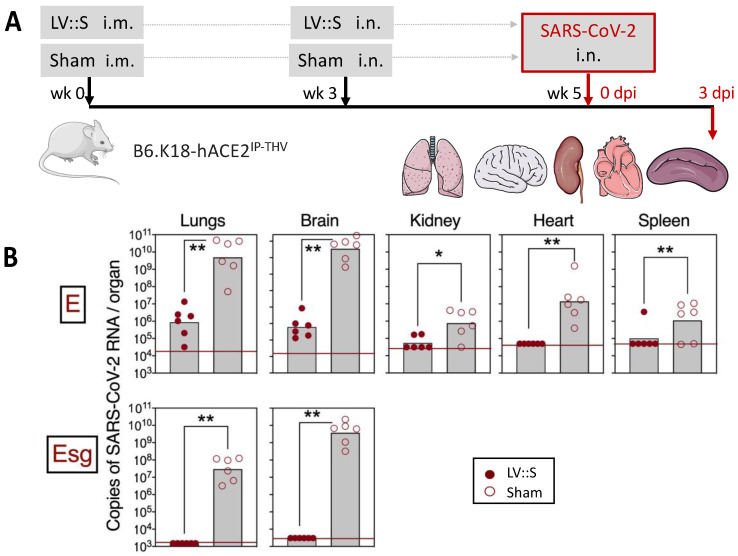
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
