

Membrane remodelling triggers maturation of excitation-contraction coupling in 3D-shaped human-induced pluripotent stem cell-derived cardiomyocytes
- PMID: 36988697
- PMCID: PMC10060306
- DOI: 10.1007/s00395-023-00984-5
Membrane remodelling triggers maturation of excitation-contraction coupling in 3D-shaped human-induced pluripotent stem cell-derived cardiomyocytes
Abstract
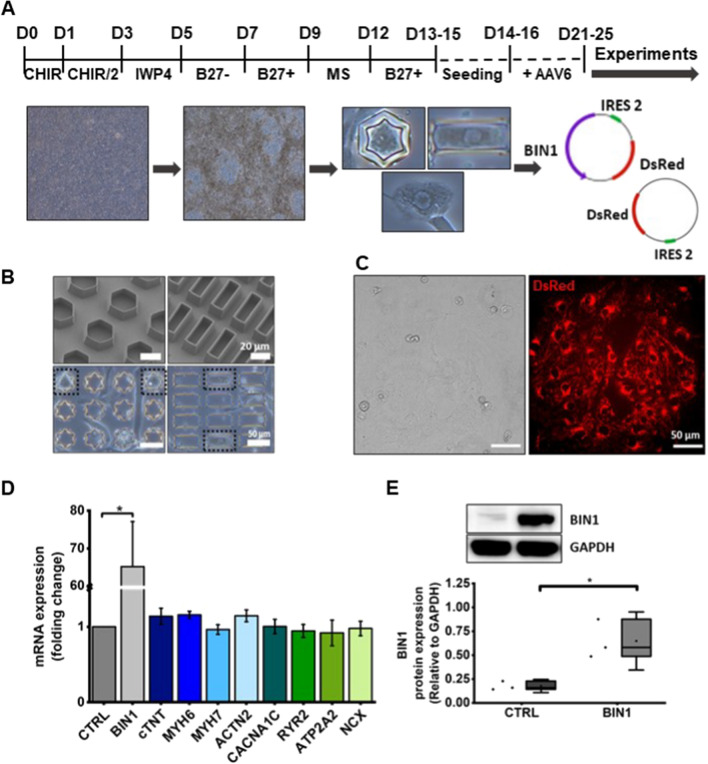
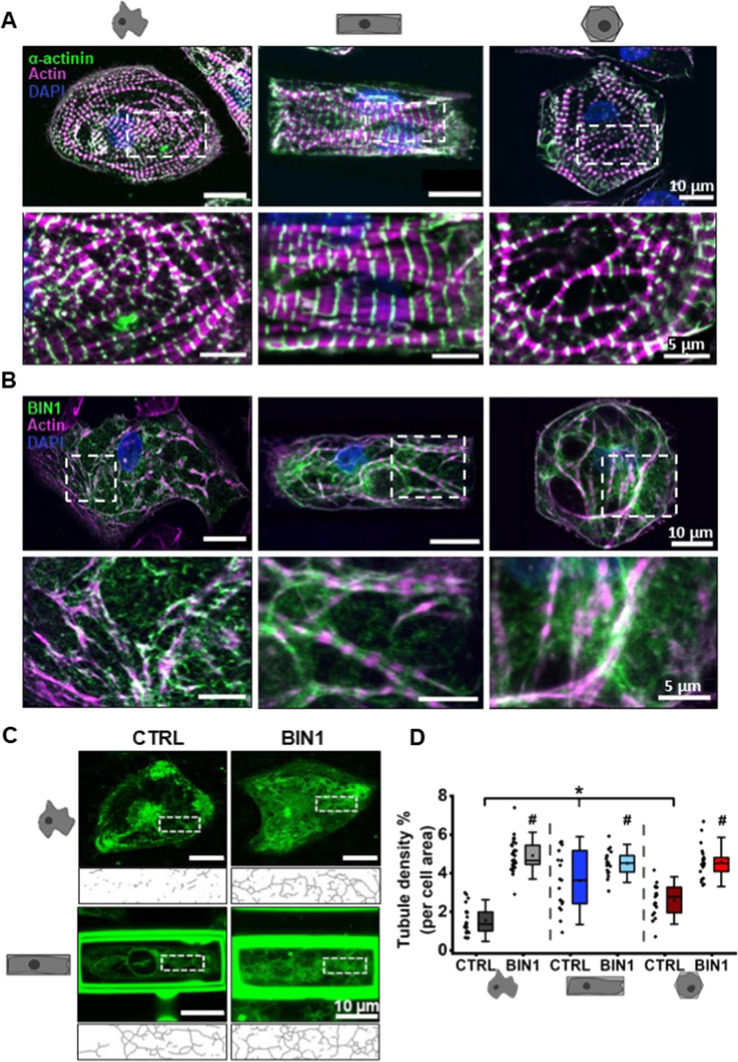
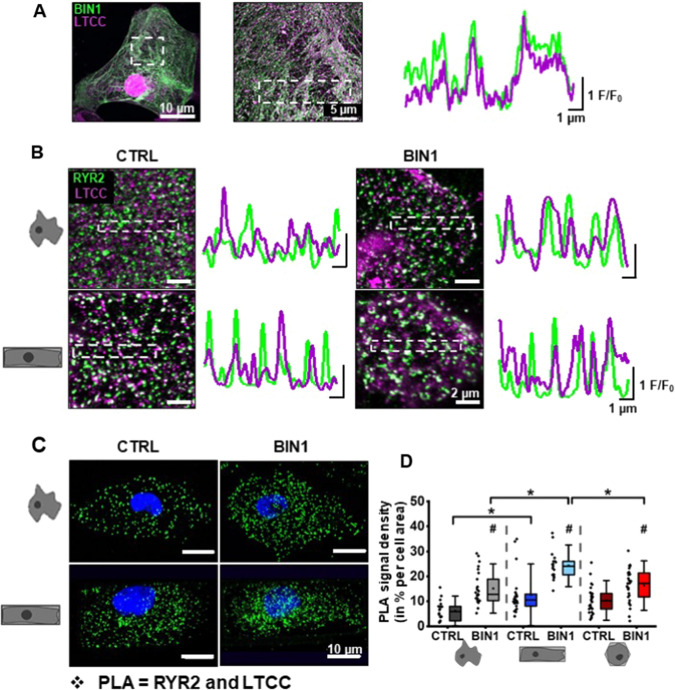
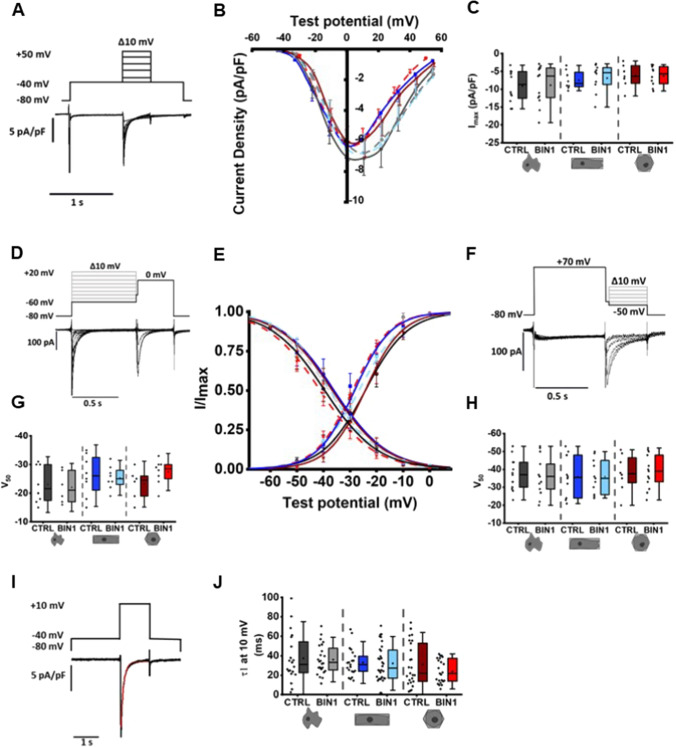
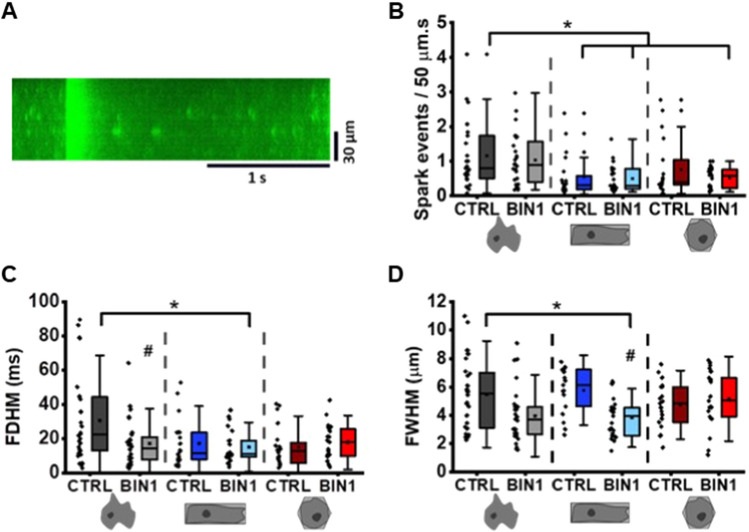
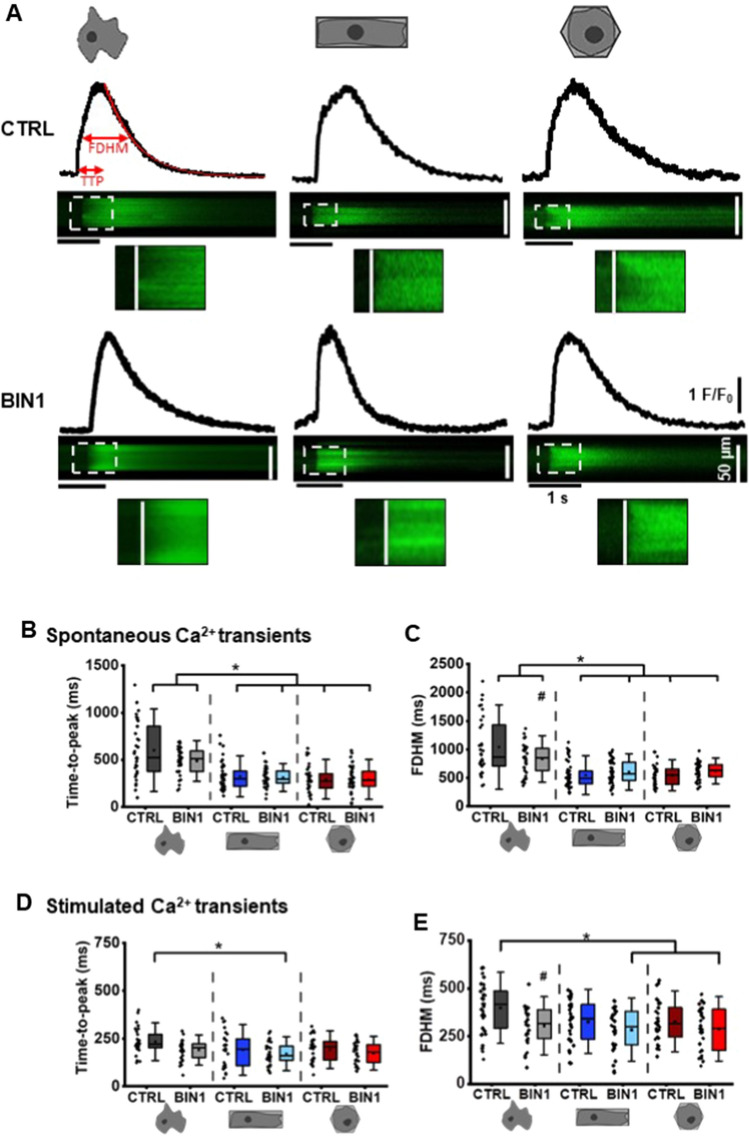
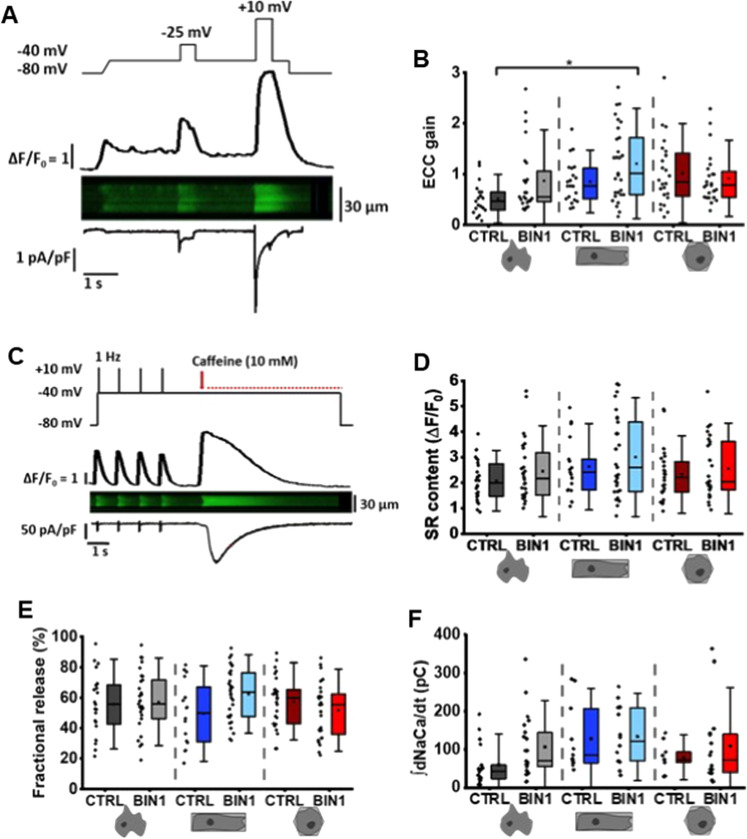
The prospective use of human-induced pluripotent stem cell-derived cardiomyocytes (hiPSC-CM) for cardiac regenerative medicine strongly depends on the electro-mechanical properties of these cells, especially regarding the Ca2+-dependent excitation-contraction (EC) coupling mechanism. Currently, the immature structural and functional features of hiPSC-CM limit the progression towards clinical applications. Here, we show that a specific microarchitecture is essential for functional maturation of hiPSC-CM. Structural remodelling towards a cuboid cell shape and induction of BIN1, a facilitator of membrane invaginations, lead to transverse (t)-tubule-like structures. This transformation brings two Ca2+ channels critical for EC coupling in close proximity, the L-type Ca2+ channel at the sarcolemma and the ryanodine receptor at the sarcoplasmic reticulum. Consequently, the Ca2+-dependent functional interaction of these channels becomes more efficient, leading to improved spatio-temporal synchronisation of Ca2+ transients and higher EC coupling gain. Thus, functional maturation of hiPSC-cardiomyocytes by optimised cell microarchitecture needs to be considered for future cardiac regenerative approaches.
Keywords: 3D reshaping; BIN1; Excitation–contraction coupling; Maturation; hiPSC cardiomyocytes; t-tubules.
© 2023. The Author(s).
Conflict of interest statement
None to declare.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
