

mRNA translational specialization by RBPMS presets the competence for cardiac commitment in hESCs
- PMID: 36989351
- PMCID: PMC10058251
- DOI: 10.1126/sciadv.ade1792
mRNA translational specialization by RBPMS presets the competence for cardiac commitment in hESCs
Abstract
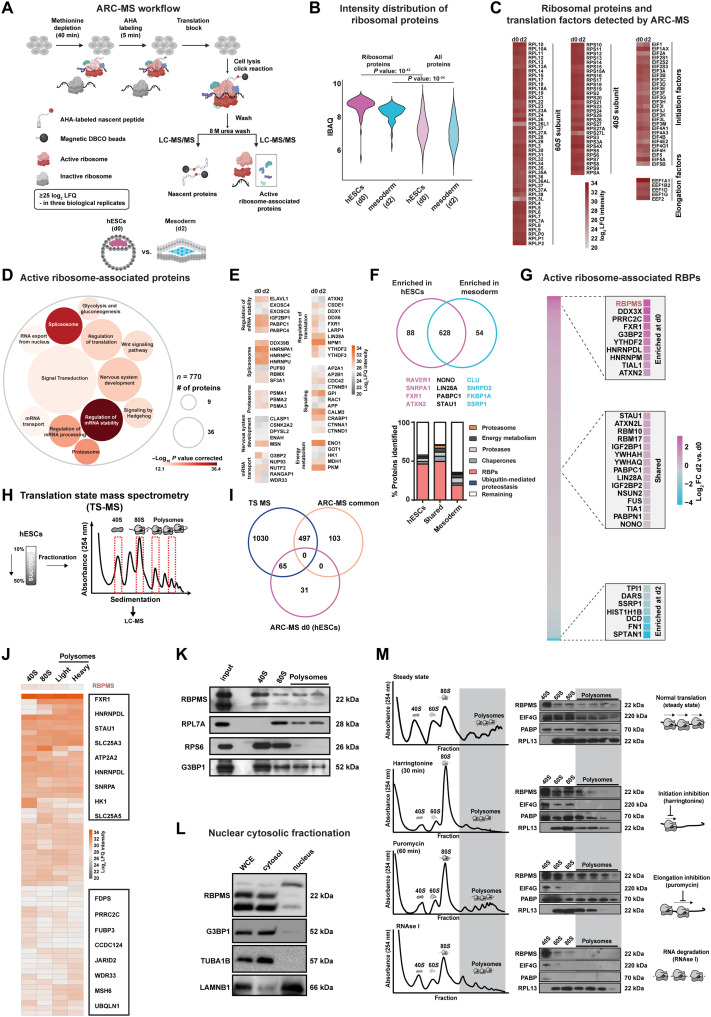
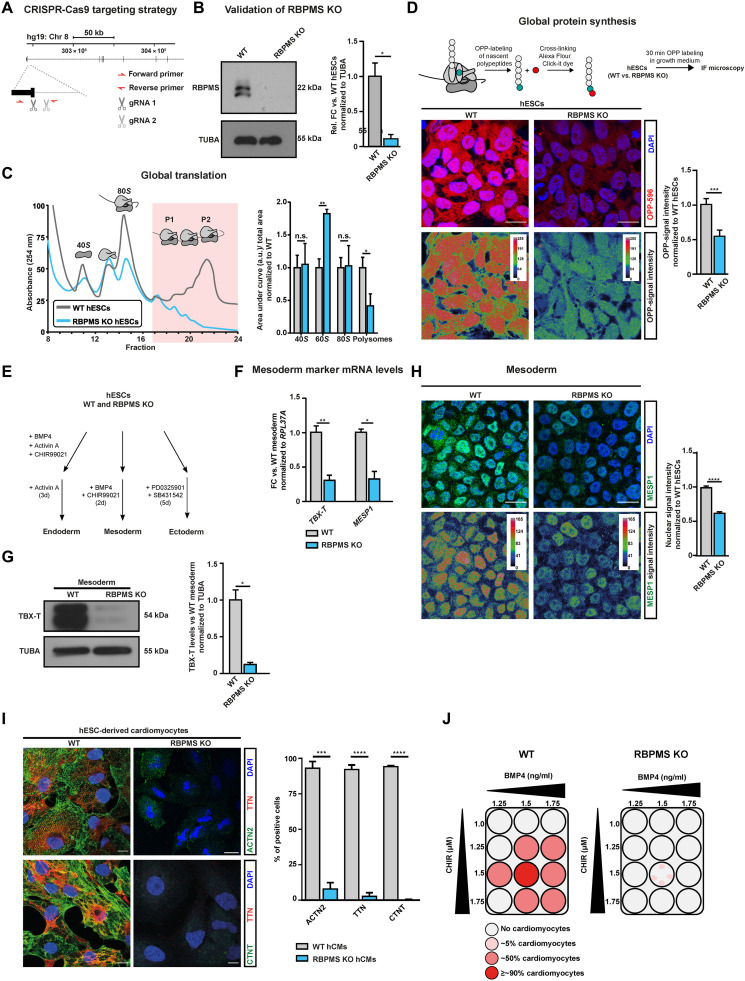
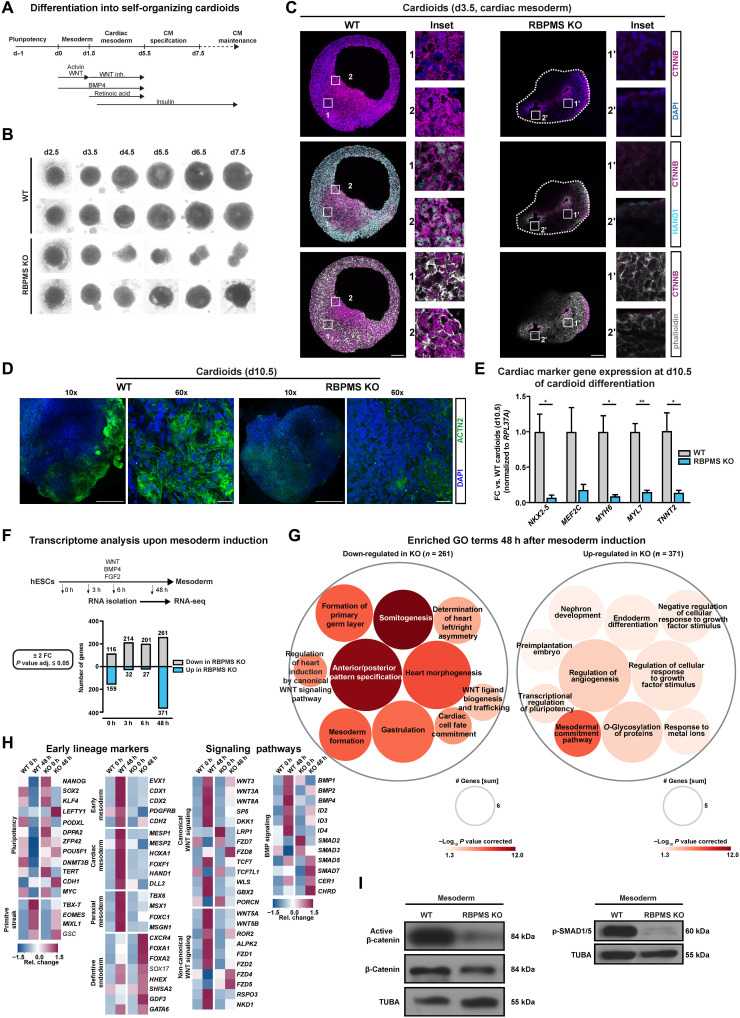
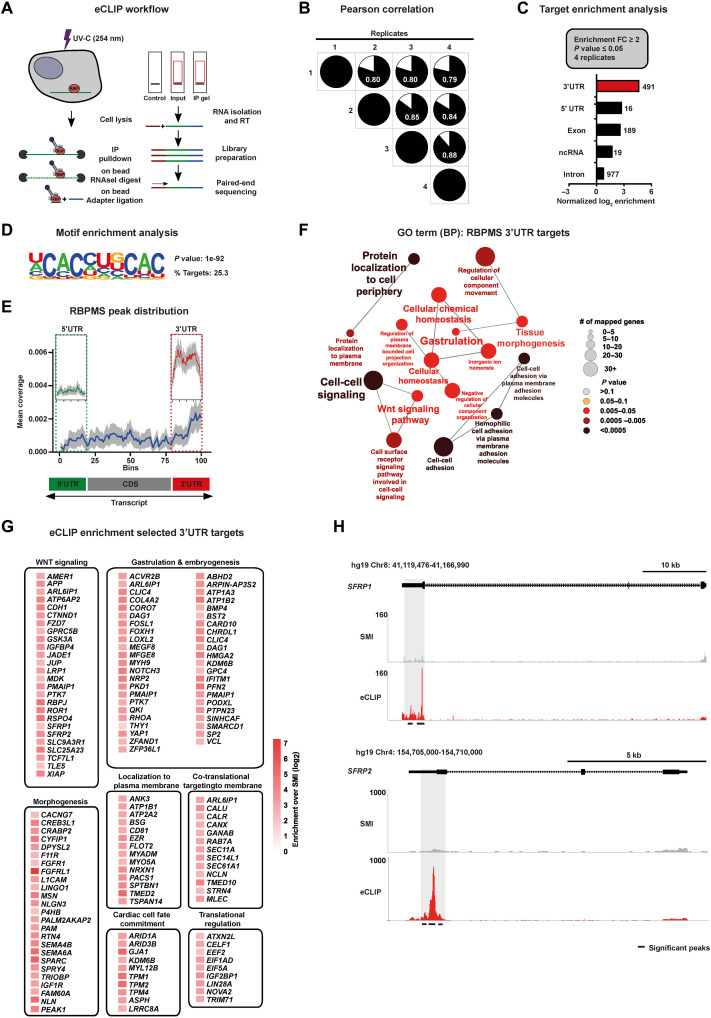
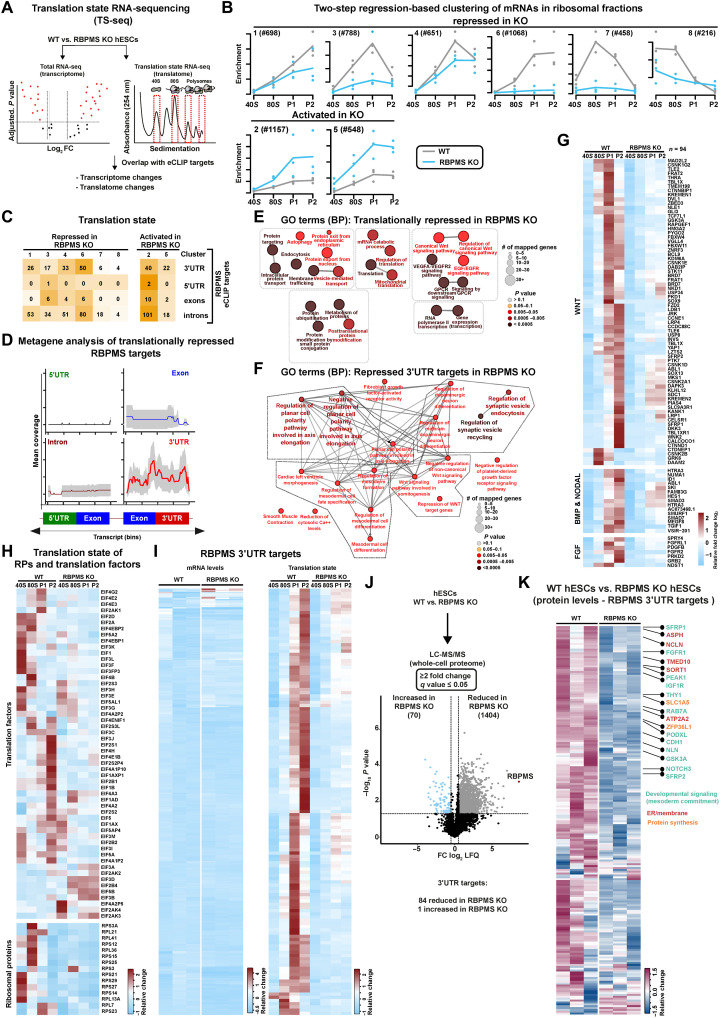
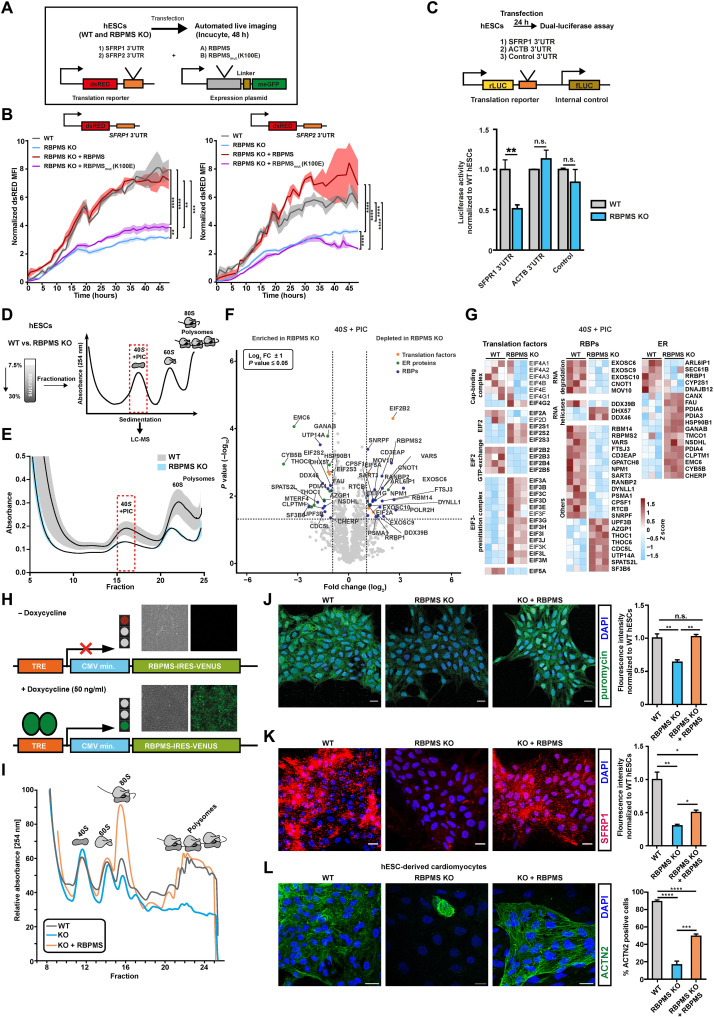
The blueprints of developing organs are preset at the early stages of embryogenesis. Transcriptional and epigenetic mechanisms are proposed to preset developmental trajectories. However, we reveal that the competence for the future cardiac fate of human embryonic stem cells (hESCs) is preset in pluripotency by a specialized mRNA translation circuit controlled by RBPMS. RBPMS is recruited to active ribosomes in hESCs to control the translation of essential factors needed for cardiac commitment program, including Wingless/Integrated (WNT) signaling. Consequently, RBPMS loss specifically and severely impedes cardiac mesoderm specification, leading to patterning and morphogenetic defects in human cardiac organoids. Mechanistically, RBPMS specializes mRNA translation, selectively via 3'UTR binding and globally by promoting translation initiation. Accordingly, RBPMS loss causes translation initiation defects highlighted by aberrant retention of the EIF3 complex and depletion of EIF5A from mRNAs, thereby abrogating ribosome recruitment. We demonstrate how future fate trajectories are programmed during embryogenesis by specialized mRNA translation.
Figures
References
-
- J. Newport, M. Kirschner, A major developmental transition in early Xenopus embryos: II. Control of the onset of transcription. Cell 30, 687–696 (1982). - PubMed
-
- J. Newport, M. Kirschner, A major developmental transition in early Xenopus embryos: I. characterization and timing of cellular changes at the midblastula stage. Cell 30, 675–686 (1982). - PubMed
-
- H. Spemann, H. Mangold, Induction of embryonic primordia by implantation of organizers from a different species. 1923. Int. J. Dev. Biol. 45, 13–38 (2001). - PubMed
-
- L. Xiang, Y. Yin, Y. Zheng, Y. Ma, Y. Li, Z. Zhao, J. Guo, Z. Ai, Y. Niu, K. Duan, J. He, S. Ren, D. Wu, Y. Bai, Z. Shang, X. Dai, W. Ji, T. Li, A developmental landscape of 3D-cultured human pre-gastrulation embryos. Nature 577, 537–542 (2020). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
