

Targeted DNA integration in human cells without double-strand breaks using CRISPR-associated transposases
- PMID: 36991112
- PMCID: PMC10620015
- DOI: 10.1038/s41587-023-01748-1
Targeted DNA integration in human cells without double-strand breaks using CRISPR-associated transposases
Abstract
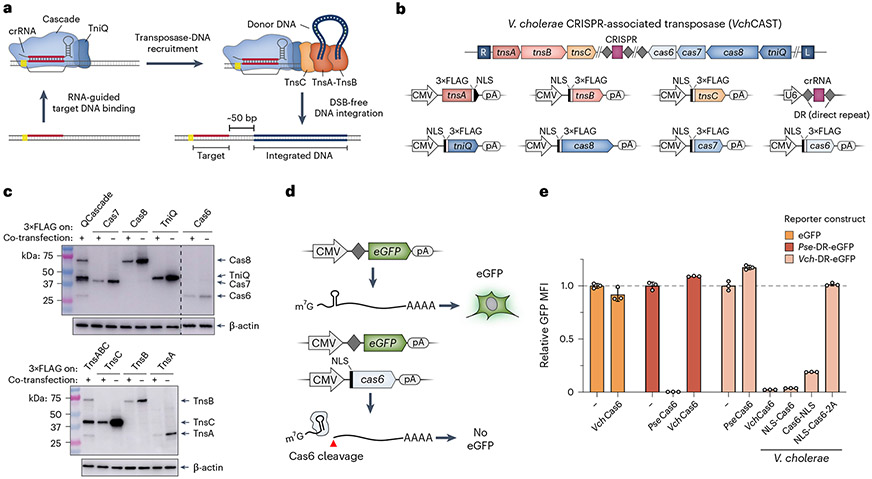
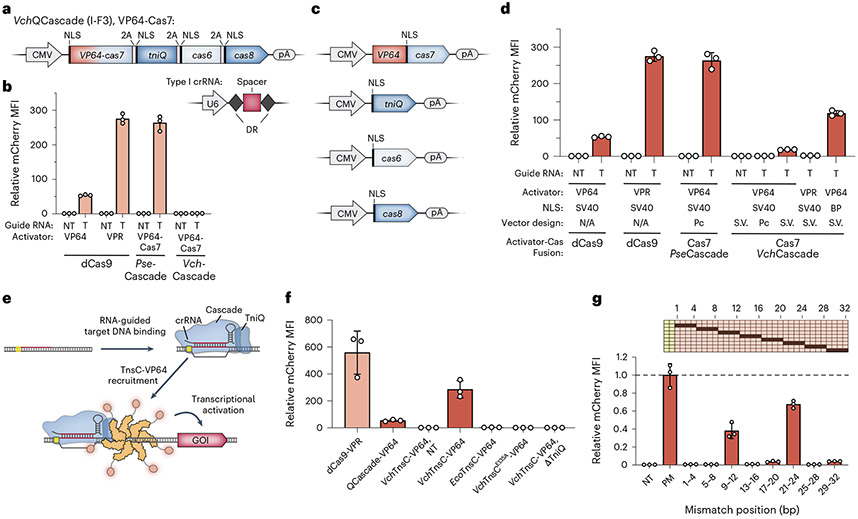
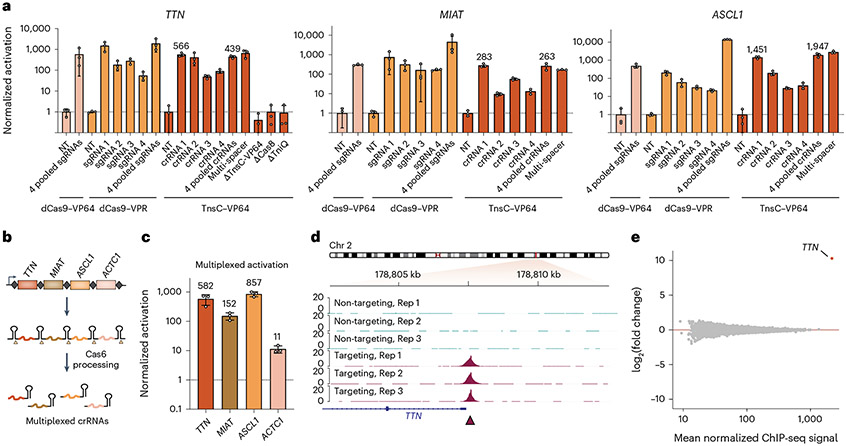
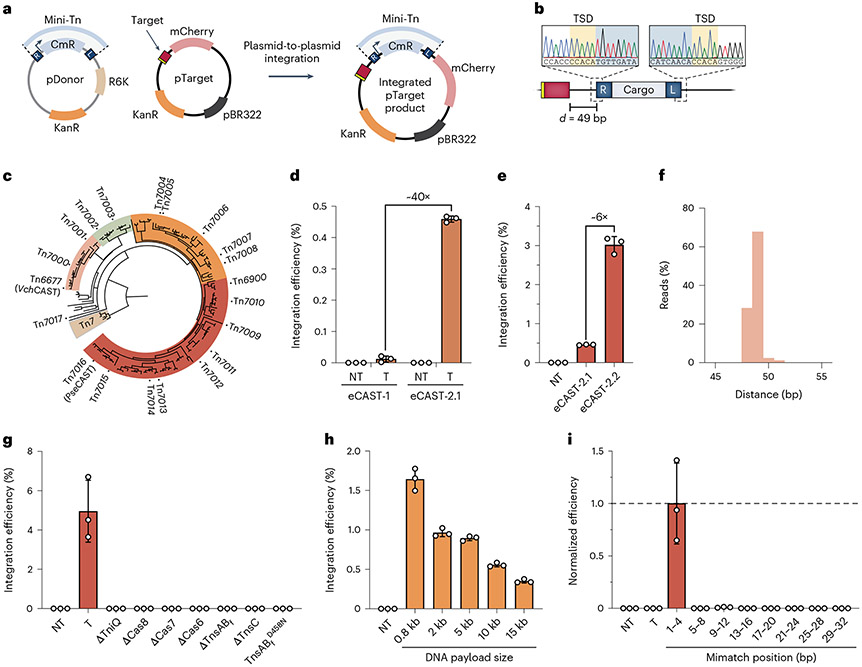
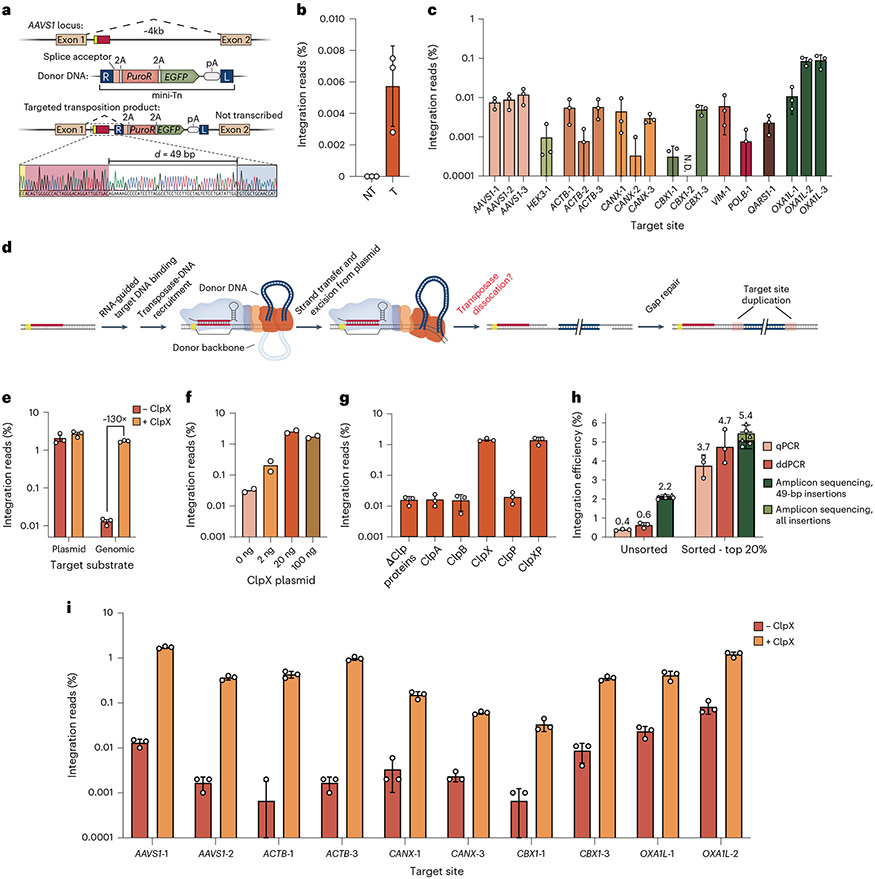
Conventional genome engineering with CRISPR-Cas9 creates double-strand breaks (DSBs) that lead to undesirable byproducts and reduce product purity. Here we report an approach for programmable integration of large DNA sequences in human cells that avoids the generation of DSBs by using Type I-F CRISPR-associated transposases (CASTs). We optimized DNA targeting by the QCascade complex through protein design and developed potent transcriptional activators by exploiting the multi-valent recruitment of the AAA+ ATPase TnsC to genomic sites targeted by QCascade. After initial detection of plasmid-based integration, we screened 15 additional CAST systems from a wide range of bacterial hosts, identified a homolog from Pseudoalteromonas that exhibits improved activity and further increased integration efficiencies. Finally, we discovered that bacterial ClpX enhances genomic integration by multiple orders of magnitude, likely by promoting active disassembly of the post-integration CAST complex, akin to its known role in Mu transposition. Our work highlights the ability to reconstitute complex, multi-component machineries in human cells and establishes a strong foundation to exploit CRISPR-associated transposases for eukaryotic genome engineering.
© 2023. The Author(s), under exclusive licence to Springer Nature America, Inc.
Figures
Update of
-
Targeted DNA integration in human cells without double-strand breaks using CRISPR RNA-guided transposases.bioRxiv [Preprint]. 2023 Mar 18:2023.03.17.533036. doi: 10.1101/2023.03.17.533036. bioRxiv. 2023. Update in: Nat Biotechnol. 2024 Jan;42(1):87-98. doi: 10.1038/s41587-023-01748-1. PMID: 36993517 Free PMC article. Updated. Preprint.
References
-
- Anzalone AV, Koblan LW & Liu DR Genome editing with CRISPR–Cas nucleases, base editors, transposases and prime editors. Nat. Biotechnol 38, 824–844 (2020). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
