

This is a preprint.
Germline-mediated immunoediting sculpts breast cancer subtypes and metastatic proclivity
- PMID: 36993286
- PMCID: PMC10055121
- DOI: 10.1101/2023.03.15.532870
Germline-mediated immunoediting sculpts breast cancer subtypes and metastatic proclivity
Update in
-
Germline-mediated immunoediting sculpts breast cancer subtypes and metastatic proclivity.Science. 2024 May 31;384(6699):eadh8697. doi: 10.1126/science.adh8697. Epub 2024 May 31. Science. 2024. PMID: 38815010
Abstract
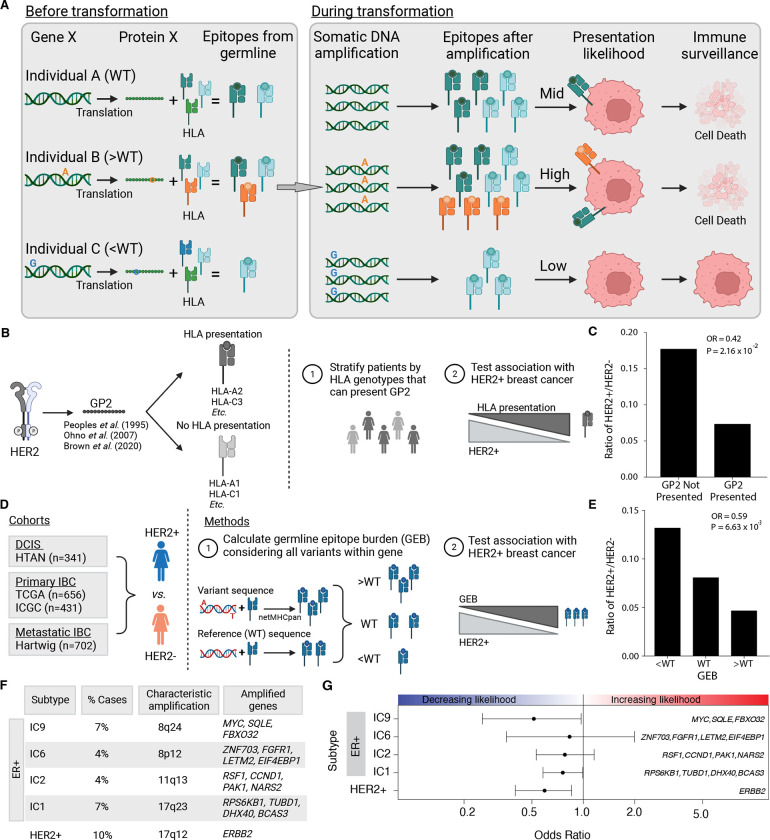
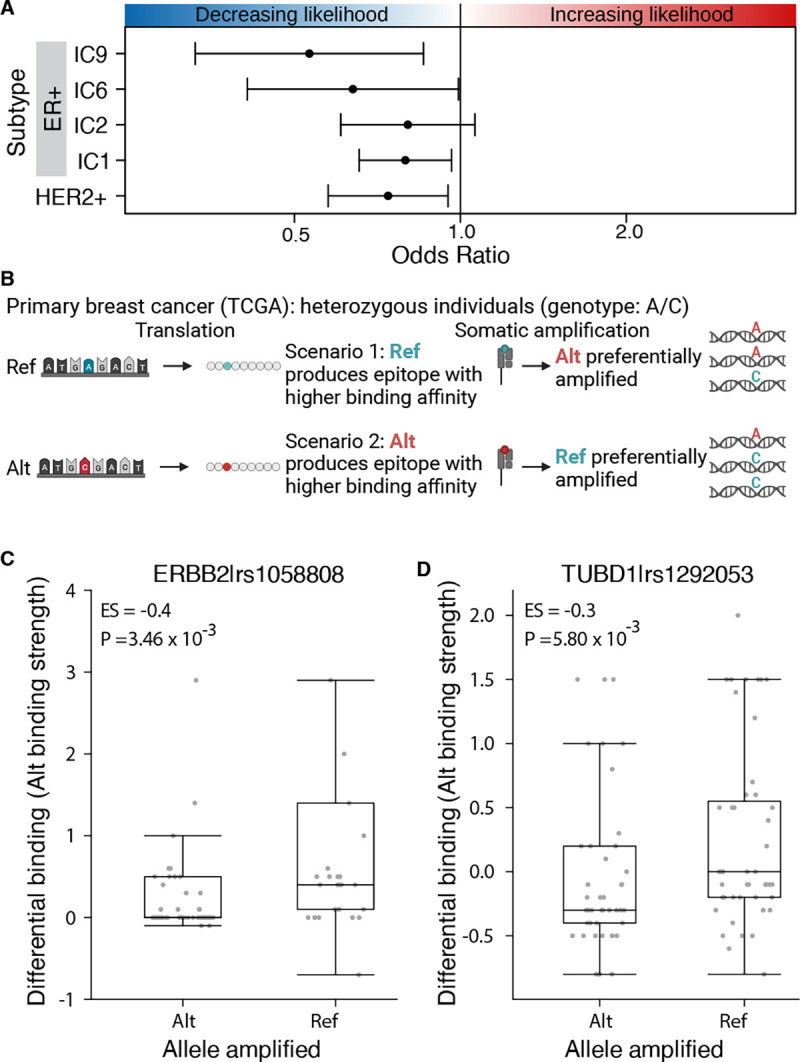
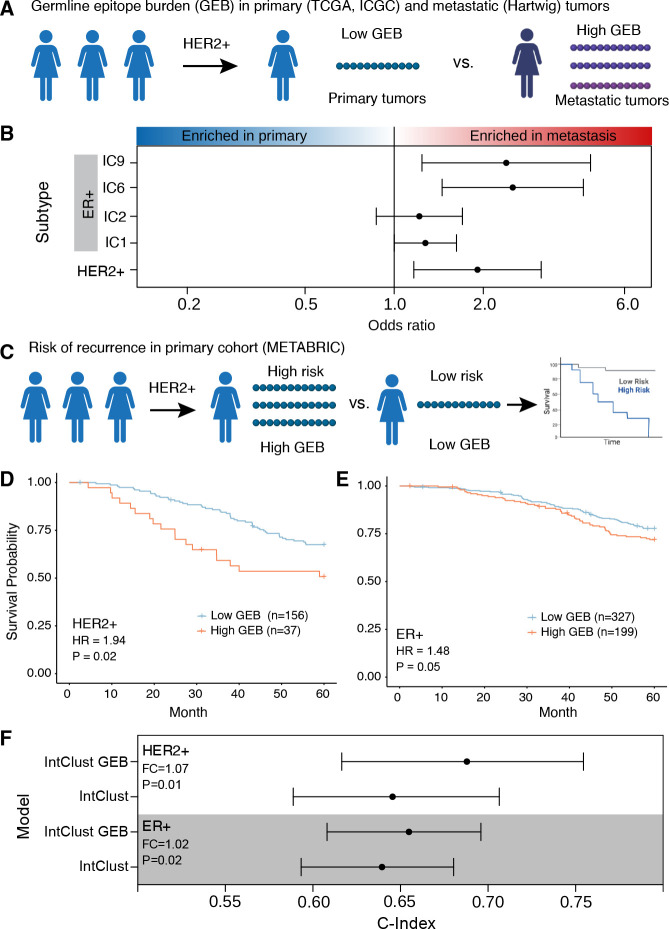
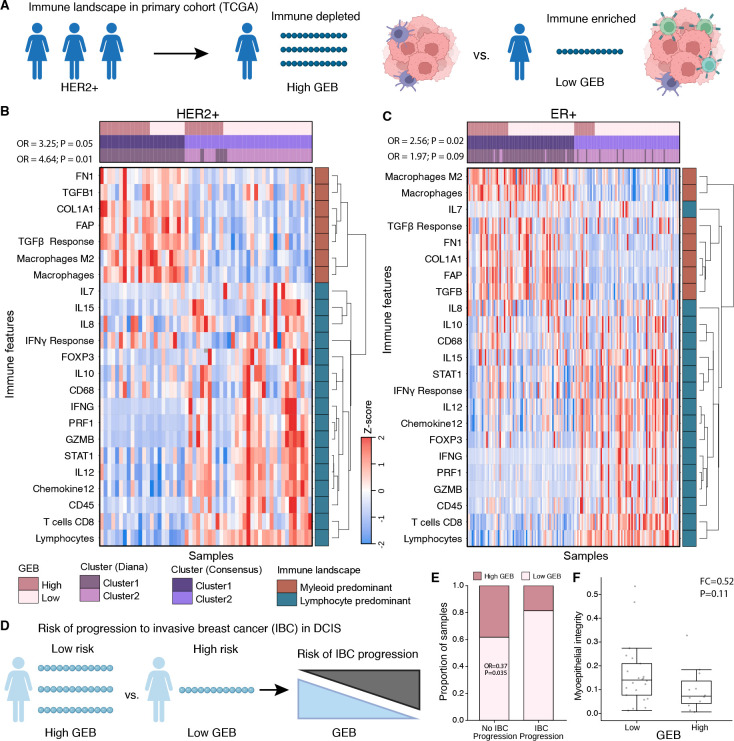
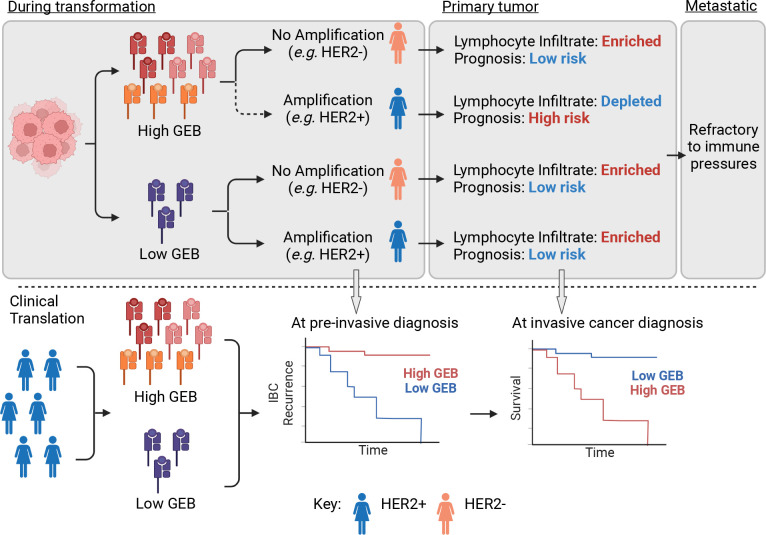
Cancer represents a broad spectrum of molecularly and morphologically diverse diseases. Individuals with the same clinical diagnosis can have tumors with drastically different molecular profiles and clinical response to treatment. It remains unclear when these differences arise during disease course and why some tumors are addicted to one oncogenic pathway over another. Somatic genomic aberrations occur within the context of an individual's germline genome, which can vary across millions of polymorphic sites. An open question is whether germline differences influence somatic tumor evolution. Interrogating 3,855 breast cancer lesions, spanning pre-invasive to metastatic disease, we demonstrate that germline variants in highly expressed and amplified genes influence somatic evolution by modulating immunoediting at early stages of tumor development. Specifically, we show that the burden of germline-derived epitopes in recurrently amplified genes selects against somatic gene amplification in breast cancer. For example, individuals with a high burden of germline-derived epitopes in ERBB2, encoding human epidermal growth factor receptor 2 (HER2), are significantly less likely to develop HER2-positive breast cancer compared to other subtypes. The same holds true for recurrent amplicons that define four subgroups of ER-positive breast cancers at high risk of distant relapse. High epitope burden in these recurrently amplified regions is associated with decreased likelihood of developing high risk ER-positive cancer. Tumors that overcome such immune-mediated negative selection are more aggressive and demonstrate an "immune cold" phenotype. These data show the germline genome plays a previously unappreciated role in dictating somatic evolution. Exploiting germline-mediated immunoediting may inform the development of biomarkers that refine risk stratification within breast cancer subtypes.
Conflict of interest statement
Conflict of Interest Statement Unrelated to this work, C.C. is an advisor to Genentech, Bristol Myers Squibb, 3T Biosciences, Resistance Bio, DeepCell, NanoString and has equity in DeepCell, Grail/Illumina.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous